Морфология растений
- Определение морфологии растений, ее задачи и методы
- Основные понятия морфологии растений. Ветвление
- Явление симметрии
- Метаморфоз, гомологии, аналогии, редукции, атавизм
- Морфология сосудистых растений
- Морфология членов сосудистых растений. Морфология корня
- Метаморфозы корней
- Корни с особыми функциями.
- Определение корня
- Морфология стебля
- Морфология листа
- Простые листья
- Сложные листья
- Метаморфозы и редукция листьев
- Морфология побега
- Специализация и метаморфоз побегов
- Жизненные формы
- Редукция вегетативных органов у цветковых паразитов и полупаразитов
- Морфология органов растения
- Стробилы и цветки
- Структурная морфология пыльцы и семязачатка
- Морфология гинецея
- Морфология соцветий
- Морфология семян
- Морфология плодов
- Морфология соплодий
- Морфологическая эволюция покрытосеменных растений
- Жизненные формы. Корневые системы
- Стебли, лист и листорасположение
- Цветок и соцветие
- Семя
- Плоды
МОРФОЛОГИЯ РАСТЕНИЙ
Определение морфологии растений, ее задачи и методы
Морфология растений изучает форму, строение, изменение структур в процессе индивидуального развития растений, формирование их в ходе филогенеза.
В морфологии растений существуют различные направления. Главными из них являются:
1) описательная морфология — занимается изучением многообразия внешнего строения растений;
2) сравнительная морфология — исследует сведения о внешнем строении растений путем сравнительного изучения;
3) эволюционная морфология — изучает внешнее строение растений с целью определения путей и направлений эволюции членов и органов растений.
Основными методами морфологии растений являются наблюдение и сравнение. Кроме того, морфология растений широко использует в своих исследованиях эксперимент.
Эксперимент в морфологии предполагает изучение ответных реакций растений на воздействие различных факторов среды. Экспериментальная морфология помогает выяснить связь между закономерностями формообразований растений и факторами внешней среды.
Важнейшими объектами морфологии растений являются органы.
Изучение развития высших растений показывает, что основными их органами являются корень, стебель и лист. Все остальные разнообразные органы произошли в результате видоизменений корня, стебля, листа. Поэтому в морфологии для этих трех органов используется термин «член».
Вся история развития морфологии растений говорит о значимости этого раздела ботаники. Через морфологию растений проявляются все признаки организма: анатомические, биохимические, генетические и др.
Морфология растений тесно связана с систематикой растений, основным методом которой является морфолого-географический метод. Морфология дает материал о происхождении и тождестве органов растений, что способствует созданию филогенетической системы растений, которая отображает родственные связи между таксонами.
Обособление таких направлений в биологии как вирусология, бактериология, микология, фикология, бриология (раздел биологии, изучающий мхи) привело к возникновению соответствующих разделов морфологии: морфология вирусов, морфология бактерий и т.д. Объектом морфологии растений, фактически, стали только сосудистые растения.
Основные понятия морфологии растений. Ветвление
Различные системы ветвления могут быть сведены к четырем типам:
1. Дихотомическое или вильчатое ветвление, — ось на вершине разделяется на две новые, дающие одинаково развитые оси.
2. Моноподиальное ветвление — главная ось не прекращает роста в длину и образует ниже своей вершины, обычно в восходящей последовательности, боковые оси.
3. Ложнодихотомическое ветвление. В этом случае рост главной оси прекращается, а под вершиной ее образуются две одинаковые оси, которые со временем перерастают главную ось.
4. Симподиальное ветвление. В одном случае одна из осей получает более сильное развитие, сдвигает в сторону боковую и принимает направление главной оси. Второй вариант связан с прекращением роста главной оси или смещением ее вбок, которую заменяет развивающаяся под верхушкой боковая ось. В дальнейшем эта ось ведет себя как главная и ее заменяет новая.
Эволюционно эти типы ветвления связаны следующим образом. Основными системами ветвления считаются дихотомическое, которое встречается у многих водорослей, некоторых грибов и мхов, у плаунов, и моноподиальное, которое бывает у водорослей, большинства грибов, мхов, хвощей, у многих семенных растений. Из моноподиального ветвления выводится ложнодихотомическое, которое встречается у сирени, каштана конского и др. Первый вариант симподиального ветвления выводится из дихотомического ветвления (селагинелла), а второй из моноподиального ветвления. Симподиальное ветвление, возникшее из моноподиального, встречается у многих хорошо известных нам растений, например, липы, ивы, березы, земляники, живучки, лютиков, клевера гибридного, у видов семейства пасленовые.
Типы ветвления, число, размеры и направления осей определяют внешний облик растения — габитус, который позволяет часто узнать вид многих растений даже визуально.
Явление симметрии
Симметрия — явление свойственное как для неживой природы, так и для растений, грибов, животных и прочих представителей живых организмов.
Если через ось растения или через какую-нибудь другую его часть можно провести три или более плоскостей симметрии, то такое строение называется полисимметричным или радиальным. Радиальную симметрию имеют стебли, корни. Околоцветник и его части (венчик, чашечка) с радиальной симметрией часто называют не радиальным, а актиноморфным («акстис» — луч, «морфе» — форма).
Если через ось растения, или через какую-нибудь другую его часть, можно провести только две плоскости симметрии, то говорят о бисимметрии или о билатеральной симметрии. Это относится к плоским стеблям кактусов опунций, листьям ириса и т.д.
Если через ось растения или через какую-нибудь его часть можно провести только одну плоскость симметрии, то такое строение называют моносимметричным. Например, моносимметричное строение имеют большинство листьев. Моносимметричные околоцветники, чашечки, венчик называют зигоморфными («зигон» — ярмо — часть повозки, куда запрягают пару волов; «морфе» — форма).
Если через ось, или другую часть растения, нельзя провести и одну плоскость симметрии, то такое строение называют асимметричным. Асимметричны листья вяза, цветки канны, валерианы.
У горизонтально расположенных частей растения, где можно выделить верхнюю и нижнюю части, которые часто отличаются по строению, окраске, говорят о дорзовентральном строении. Дорзовентральными являются листья большинства высших растений.
Метаморфоз, гомологии, аналогии, редукции, атавизм
Путем сравнительно-морфологического изучения все многообразие форм тела высших растений можно свести к трем основным членам: корню, стеблю, листу. У многих растений в процессе эволюции те или иные части сильно видоизменены, подверглись метаморфозу (превращению). Эти метаморфозы наследственно закреплены. Примерами метаморфозов являются усики гороха, усы винограда, колючки, корнеплоды и т.п. Однотипные метаморфозы могут иметь различное происхождение. Так, усики гороха — видоизменение листочков, а усы винограда — видоизменение соцветий.
Органы растения или их части, имеющие общее происхождение, но отличающиеся по внешнему строению, называют гомологами (согласие).
Гомологичны, например, лист, колючка барбариса, ловчий аппарат насекомых у росянки.
Органы или части их, выполняющие одинаковую функцию и приспособленные к ней, имеющие одинаковый внешний вид, но различающиеся по своему морфологическому происхождению от разных исходных форм, называются аналогичными (соответствие). Аналогичны, например, колючки барбариса, боярышника, гледичии, т.к. колючка барбариса имеет листовое происхождение, а колючка боярышника и гледичии — стеблевое.
У многих растений встречаются наследственно закрепленные редукции («редукцио» — возвращение назад) тех или иных органов или их частей. Редуцированы, например, листья у паразитных растений, редуцирован главный корень у многих корневищных растений, редуцирована чашечка у некоторых видов зонтичных и у всех видов сложноцветных. У некоторых растений наблюдается и полная утрата, или абортирование, того или иного органа или его частей. Например, у вероники произошло абортирование 3 тычинок, у крестоцветных — исчезли кроющие листья. Иногда у растений наблюдаются признаки, не свойственные данному виду, но имевшиеся у отдаленных предков. Это атавизм («атавизм» — отдаленный предок). Например, у первоцветных известны особи с 10 тычинками, проростки картофеля имеют цельные листья и т.д.
Чрезвычайно широко распространено у растений явление корреляции — зависимости развития одного органа от другого. Например, увеличение размеров колоса приводит к уменьшению размера стебля. Нарушение целостности корня приводит к увеличению количества боковых корней, ранение стебля приводит к образованию придаточных почек и т.д.
Морфология сосудистых растений
Морфология сосудистых растений — это морфология членов и органов. Членами сосудистых растений являются корень, стебель, лист и их видоизменения. К органам сосудистых растений относятся стробилы, цветки, семена и плоды.
Морфология членов сосудистых растений. Морфология корня
Корень в процессе эволюции возник как член растения, обеспечивающий его существование на суше. Основными функциями корня, с момента его образования, стали — удержание растения в почве и обеспечение растения водой и питательными солями.
В период эволюционного развития корни растений приобрели и другие функции: накопление питательных веществ, первичный синтез некоторых органических соединений, микоризообразование, образование придаточных почек и пр.
Корень, развивающийся из зародышевого (первичного) корня, называется главным. Участок на границе между главным корнем и стеблем называется корневой шейкой. В этом месте четко заметен переход от стебля к корню. Выше корневой шейки расположено подсемядольное колено стебля, или гипокотиль («гипо» — внизу, «котиледон» — семядоля) — часть стебля между корневой шейкой и семядолями. В этой части стебля эндогенно образуются, так называемые, придаточные корни. Этот вид корней может образовываться и в районе узлов надземных и подземных побегов, а иногда и в районе междоузлий надземных побегов.
Кроме главного и придаточных корней у большинства растений образуются боковые корни. Возникают они эндогенно на главном и придаточных корнях. В дальнейшем возможно образование боковых корней последующих порядков, которые формируют уже систему боковых корней.
По форме большинство корней бывают нитевидными или веретеновидными. Кроме того, корни бывают конусовидные, реповидные, клубневидные. Окраска корней варьирует от желтой до бурой, может быть даже зеленой.
К морфологическим признакам относится расчленение корня на зоны. Определение границ зон и их название относится к дискуссионным вопросам. С точки зрения морфологии различают три зоны: I) зона роста — от вершины корня до корневых волосков; 2) зона корневых волосков — часть корня, где развиваются корневые волоски; 3) зона проведения -часть корня от зоны корневых волосков до стебля.
На вершине корня расположен корневой чехлик, считающийся одним из отличительных признаков корня.
Совершенство функций корня привело к созданию в процессе эволюции корневой системы — совокупности всех корней растения. Морфологи различают три типа корневых системы:
I. Система главного корня. Состоит из главного и боковых корней. Бывает стержневой или ветвистой. В первом случае главный корень растет в длину весь период онтогенеза. Во втором случае на ранних этапах онтогенеза главный корень начинает ветвиться за счет раздвоения верхушки.
II. Система придаточных корней. Образована придаточными и боковыми корнями, редко только придаточными. Бывает мочковатой, когда придаточные корни связаны с гипокотилем или пучковатой, когда придаточные корни образуются в районе узлов надземных или подземных побегов.
III. Смешанная корневая система. Образована главным, придаточными и боковыми корнями.
Следует сказать, что в период онтогенеза тип корневой системы может меняться. На ранних этапах индивидуального развития, растение может иметь систему главного корня, потом формируется смешанная корневая система, а после отмирания главного корня растение будет иметь систему придаточных корней. Такое явление характерно для корневищных двудольных растений.
От мощности развития корневой системы зависит жизнедеятельность растения. Под землей у растений находится такая же, а может и большая масса их тела, чем над землей.
Общая длина всех корней отдельных видов культурных злаков превышает 500 м, а общая длина корней тыквы — около 25 км.
Рекомендации по внедрению растений в культуру должны сопровождаться данными о глубине и ширине залегания корневой системы. Исследования в этом направлении дают интересные цифры. У известной верблюжьей колючки корни проникают на глубину до 20 м. Корневая система у лука репчатого достигает 60 см в диаметре, а проникает на глубину до 1м. У моркови — свыше 2 м в глубину и до 1,5 м в диаметре. При раскопе корневых систем клеверов были получены в среднем такие данные, как глубина 1-1,5 м, диаметр — 0,8-3 м.
Следует отметить, что важным морфологическим показателем является площадь поверхности корней, образующих корневую, систему. Например, у пшеницы она равняется, в среднем, 4 м2, у овса — 1,5 м2.
Морфология корней зависит от способности их вступать в симбиоз с азотфиксирующими бактериями и образовывать микоризу. В первом случае на корнях образуются утолщения — клубеньки. В случае поселения грибов на корнях, боковые корни, на которых образовалась микориза, прекращают рост и начинают ветвиться, образуя коралловидные скученные разветвления. У этих боковых корней не образуются корневой чехлик и корневые волоски.
На корнях многих двудольных образуются эндогенно придаточные почки, которые развиваются в дальнейшем в надземные облиственные побеги. Этот вариант встречается у тополя, вишни, барбариса, осота, одуванчика и др.
Форма корневой системы связана с геотропизмом корней. Главный корень углубляется в почву в отвесном направлении, образуя со стеблем одну линию. Это явление носит название положительного геотропизма. Боковые корни и придаточные растут под влиянием той же силы, что определяет положительный геотропизм — силы земного притяжения, но растут они либо параллельно поверхности почвы, либо под определенным углом. Это объясняется тем, что они поперечно геотропичны. Боковые корни третьего порядка уже растут во всех направлениях, т.к. геотропичностью не обладают.
Метаморфозы корней
Корни с особыми функциями.
В процессе эволюции корни отдельных растений приобретали дополнительные функции. Одни из них стали резко отличаться по внешнему виду от типичных корней — это и есть метаморфозы корней. Другие сохранили типичное строение. Это корни с особыми функциями.
К метаморфозам корней относятся мясистые корни — корнеплоды и корнеклубни. Корнеклубни — это видоизменения придаточных корней. Корнеклубни на вершине несут придаточные почки. Корнеклубни образуются у георгин, чистяка, земляных орешков, бататов.
Корнеплоды известны у моркови, петрушки, редьки, свеклы и др. Но следует помнить, что «корнеплод» у них в морфологическом смысле представлен укороченным стеблем — частью несущей листья, подсемядольным коленом и, наконец, собственно корнеплодом — видоизмененным мясистым корнем.
Из корней с особыми функциями можно назвать ходульные корни, служащие для опоры деревьев; корни-прицепки; воздушные корни, способные поглощать воду из атмосферы; дыхательные корни, которые поглощают воздух; зеленые ассимилирующие корни, содержащие в клетках хлоропласты.
Специализация корней с особыми функциями связана с анатомическими и физиологическими особенностями.
Определение корня
Учитывая все вышеизложенное, можно вывести морфологическое определение корня.
Корень — член растения, не несущий ни листьев, ни экзогенных почек, он может возникать на всех членах растения; имеет верхушечный рост, в большинстве случаев продолжающийся долго, ветвится эндогенно, на веркушке имеет чехлик, главный корень является положительно геотропичным.
Морфология стебля
Стебель более разнообразен по внешнему строению, чем корень. Это связано, прежде всего, с разнообразием мест обитания растений. Морфологически стебель можно определить как член растения, имеющий радиальное строение, верхушечный рост и образующий в определенном порядке листья и почки. Основные функции типичных надземных стеблей: обеспечение увеличения поверхности растений путем роста и ветвления; образование листьев и формирование листовой мозаики; обеспечение связи между корнями и листьями; образование цветков, посредством которых идет половое размножение растений. Часто в стеблях древесных растений и в стеблях подземных откладываются запасные питательные вещества.
Морфологическое разнообразие стеблей связано с их формой, размерами, окраской, наличием и особенностью опушения. У древесных стеблей добавляется характер поверхности, наличие, форма, количество чечевичек. У травянистых — положение в пространстве.
Форма стебля у большинства растений цилиндрическая, но бывает трехгранная, четырехгранная, многогранная, ребристая, сплюснутая или плоская, крылатая, бочонковидная и т.д.
Размер стебля у отдельных видов является наследственно закрепленным признаком, колебания которого возможны в пределах нормы реакции. Максимальной для растений высоты достигают эвкалипты — 145-150 м. Из наших деревьев: ель достигает 50 м высоты, сосна — 40-50 м, дуб — 40 м, береза — 30 м. Если говорить о длине стебля, то рекорд принадлежит тропическим лазающим пальмам-ратангам, длина стеблей у которых достигает 300 м. Толщина стеблей может достигать 10-12 м у баобаба и секвой. Наименьшими размерами характеризуется стебель ряски, вольфии, бульбофиллиума. Так, у вольфии он равен 1-1,5 мм, у бульбофиллиума — 2 мм, у ряски до 10 мм.
Окраска стебля травянистых растений обычно зеленая. Это связано с наличием в клетках коры этих растений хлоропластов. Довольно обычным явлением считается наличие пятен на поверхности травянистых стеблей. К концу вегетации стебель становится желтым, бурым, коричневым. Встречаются у травянистых растений и альбиносы. У древесных растений окраска молодых стеблей определяется наличием хлоропластов в клетках коры. В дальнейшем она связана с окраской пробки, покрывающей стебель. А окраска пробки варьирует от почти белой у березы до темнокоричневой.
Стебли травянистых растений могут быть голые, гладкие, а могут быть опушенные и даже покрыты шипами (розы). Опушение стеблей характеризуется интенсивностью опушения (слабо опушенные, сильно опушенные), длиной волосков (коротко опушенные, длинно опушенные); положением волосков (прижато опушенные, опушенные оттопыренными волосками); видом трихом (опушение железистое, опушение из простых волосков, опушение из звездчатых волосков, опушение из двураздельных волосков и т.д.). Стебли древесных растений бывают гладкие, чешуйчатые, ребристые, морщинистые.
Неотъемлемым элементом покровной ткани древесных растений являются чечевички, через которые осуществляется газообмен. Чечевичек может быть много или мало. Бывают они круглыми, овальными или вытянутыми.
Положение стебля в пространстве есть адаптивный морфологический признак. Различают стебли прямостоячие, приподнимающиеся, стелющиеся, вьющиеся, цепляющиеся.
Кроме того, при исследовании растений, можно обнаружить еще много важных черт морфологического строения стебля. Это и лиственные рубцы со своими пучковыми рубцами, группы терминальных рубцов почечных чешуй у древесных растений; различного рода железки у травянистых растений и т.д.
Различные взгляды на понятие «ветвление», «клубни», «корневища», приводят к тому, что морфология этих структур рассматривается на уровне стеблей или на уровне побегов. Мы считаем, что логичнее этот материал рассматривать при описании побега.
Морфология листа
Лист является самым полиморфным членом тела растения, а поэтому в определении листа морфологические особенности его не учитываются. Лист — это член растения, выполняющий в большинстве случаев функции фотосинтеза, транспирации и газообмена.
Основатель научной ботаники К.Линней насчитывал 170 различных типов листьев.
Сейчас рассматривают 2 типа листьев — простые и сложные. Все остальное многообразие листьев определяется его морфами или просто видами.
Простые листья
Эволюционно простые листья считаются первичными. Простые листья представляют собой следующие комбинации частей простого листа:
- I) листовая пластинка, черешок, прилистники;
- 2) листовая пластинка, черешок;
- 3) листовая пластинка;
- 4) листовая пластинка, черешок, влагалище;
- 5) листовая пластинка, влагалище;
- 6) листовая пластинка, язычок, влагалище;
- 7) листовая пластинка, черешок, раструб.
Листья I, 2, 4, 7 называют черешковыми, листья 3 — сидячими, листья 5, 6 — влагалищными; лист 7 -лист с раструбом, лист I — лист с прилистниками.
Наибольшее количество морфологической информации несет листовая пластинка.
Листовые пластинки морфологи описывают по их общей форме, по консистенции, по очертанию (не путать с формой) всей пластинки, по форме вершины и основания (не путать с основанием листа), по расчлененности, опушению, характеру поверхности, строению края листовой пластинки, жилкованию и т.д. В частности, широкое использование получил такой признак, как площадь листовой пластинки.
Много морфологически значимых признаков несет черешок. Главные из них — форма черешка на поперечном сечении. Чаще она бывает округлой, но может быть ребристой, выемчатой, желобчатой, плоской и т.д., а иногда и не поддающейся словесному описанию.
Прилистники могут иметь различную форму, консистенцию, размеры, окраску и т.д. Глобальным признаком считается наличие или отсутствие прилистников, хотя их функция так до конца и не понята.
Большое морфологическое значение имеет форма и строение листового влагалища. Особо значимы морфологические признаки влагалища для описания видов злаков и осок. По-своему интересен с точки зрения морфологии язычок, расположенный у листьев злаков между пластинкой и влагалищем. Он может иметь различную форму и различные размеры, признаки его являются видовыми.
Сложные листья
Встречаются у эволюционно продвинутых таксонов. Для такого семейства, как бобовые — это вообще характерный вариант типа листа. В пределах этого семейства встречаются почти все варианты сложных листьев: тройчатосложный лист — состоит из черешка, прилистников и трех листочков, расположенных на вершине черешка (клевер, люцерна, пажитник, донник); пальчатосложный лист — состоит из черешка, прилистников, четырех и более листочков на вершине черешка (клевер люпиновый, люпин); перистосложный лист — состоит из черешка, прилистников, рахиса и расположенных на нем листочков (астрагал, эспарцет, солодка), или рахиса и листочков (дориникум, лядвенец); дважды-перистосложные (гледичия, мимоза, акация).
Таким образом, постоянным элементом сложного листа являются листочки.
Сами листочки несут массу морфологической информации, сравнимой с информацией о строении простых листьев. Отсутствуют у листочков лишь прилистники и влагалище.
Особым видом листьев следует считать семядоли — первые листья зародыша. Число семядолей — характерный признак для крупных групп семенных растений: у однодольных — одна семядоля; у двудольных обычно две семядоли, у голосеменных — от двух до 15. Форма и размеры семядолей отличают их уже на эмбриональном уровне. У большинства растений они при прорастании семени выносятся над поверхностью почвы, увеличиваются в размерах, становятся зелеными и являются первыми листьями проростка. Отличаются эти листья от настоящих меньшими размерами, простотой формы. Они всегда цельные и без прилистников.
Размеры листьев весьма разнообразны: от нескольких миллиметров до 10-15, даже до 20 м длины у пальм рафий. Продолжительность существования листьев колеблется от одного вегетационного периода до 2-12 лет у голосеменных. У так называемых вечнозеленых двудольных листья живут до 3-4 лет (брусника, плющ, олеандр, лавр).
Поразительным долголетием отличаются листья вельвиччии. Они живут до 100 лет. Правда, при постепенном отмирании на вершине и нарастании у основания.
Метаморфозы и редукция листьев
Метаморфозы листьев не являются редким явлением. Они могут полностью (барбарис) или частично превращаться в колючки (чертополох), в усики (горох, вика, чина). Довольно часто листья в процессе эволюции видоизменяются в чешуйки. У луковиц листья превратились в сочные и сухие чешуи.
У многих австралийских акаций пластинка листа не развивается, а черешок превратился в плоское образование, называемое филлодием, которое и выполняет основные функции листа. Это уже пример редукции части листа. И этой редукции чаще всего подвергается листовая пластинка. У солероса от листьев остаются лишь влагалища, у черного саксаула — основания черешков.
Особую морфу представляют листья насекомоядных растений. Это и захлопывающиеся листья, и листья в виде кувшинов, и розетко-образующие листья.
В заключение попробуем все-таки дать морфологическое определение листа. Лист — это надземный член тела растения, никогда не образующий на себе другого листа.
Морфология побега
Побегом называют комплекс, состоящий из стебля и листьев. Первичный побег закладывается в зародыше, где он представлен почечкой. Почечка состоит из зародышевого стебелька — эпикотиля, апикальной меристемы и одного или нескольких листовых примордиев (зачатков листьев). При прорастании семени стебелек удлиняется. Из апикальной меристемы развиваются новые листовые примордии, из листовых примордиев развиваются листья, а в пазухах листьев формируются примордии почек. Этот алгоритм развития в ходе образования побеговой системы растения многократно может повторяться.
У сформировавшегося побега различают узлы — часть побега, где лист соединяется со стеблем; междоузлия — часть побега между узлами, обычно это часть стебля; листовые пазухи — угол между листом и восходящей частью стебля.
Частью побега являются и почки. Это, прежде всего, верхушечная почка, представляющая конус нарастания побега. В пазухах листьев у семенных растений образуются пазушные, или боковые почки. Если они развиваются одна над другой (жимолость, грецкий орех, робиния и др.), то называются сериальными. Если почки развиваются в пазухах листьев рядом, друг около друга (слива, злаки и пр.), то они называются коллатеральными. Почки могут образовываться эндогенно в районе междоузлий. Эти почки называются придаточными.
У деревьев и кустарников холодного и умеренного климата образуются зимующие, или покоящиеся почки, которые часто называют глазками. Из этих почек на следующий год развиваются новые побеги. Наружные листья у этих почек обычно превращаются в почечные чешуи, защищающие внутренние части почки от повреждения.
Зимующие, или покоящиеся почки образуются и у многолетних трав, на тех органах, которые не отмирают на зиму, т.е. на корневищах, у основания стеблей и т.п. Эти почки называют почками возобновления. Из них весной развиваются надземные побеги.
Все вышеперечисленные почки называются вегетативными. Такие почки состоят из апекса, зачаточных узлов, зачаточных междоузлий, листовых примордиев, выше которых могут развиваться почечные примордии, и зачаточных листьев.
Из почки, не имеющей почечных примордиев, развивается простой или неветвистый побег. Из почки с почечными примордиями развивается ветвистый побег.
Кроме того, у семенных растений бывают еще и генеративные почки. Это цветковые почки и почки, дающие начало шишкам голосеменных. Они отличаются от вегетативных по внешнему виду. Кроме апекса, зачаточных междоузлий и зачаточных узлов, такие почки имеют примордии, которые дают начало частям цветка или частям шишек. У почек, дающих начало соцветиям, образуются цветковые примордии.
Наконец, есть, так называемые, смешанные почки, из которых образуются облиственные побеги с цветками.
Морфологическая характеристика побега подразумевает описание строения узлов, междоузлий, почек. Обязательно указывается тип листорасположения. У большинства растений он является очередным — в узле расположен один лист, но может быть супротивным или мутовчатым. Определенный тип листорасположения формирует листовую мозаику, что позволяет наилучшим образом использовать пространство для обеспечения равномерного освещения листа.
С процессом роста и развития побега связано и деление листьев на три категории: низовые листья, срединные листья, верхушечные, или верховые листья. При морфологическом описании листьев обычно описывают срединные листья, но полное морфологическое описание требует отдельного описания всех категорий листьев, т.к. даже срединные листья на одном побеге имеют различия. Это явление носит название гетерофиллии или разнолистности.
Специализация и метаморфоз побегов
В процессе эволюции растений происходило адаптивное развитие побега в целом. Появились побеги удлиненные и укороченные. Зачастую даже простой побег в верхней части бывает удлиненный, а в нижней — укороченный и наоборот. Это явление зависит от размеров междоузлий. Так, нижняя часть побега пастушьей сумки, типичный пример укороченного побега. А вот, так называемые, «плодушки» у яблонь — это целиком укороченный побег. К специализированным побегам можно отнести ствол и сучья, из которых формируется система скелетных осей.
Весьма обычными метаморфозами побегов являются корневища, луковицы, клубни — системы, где четко просматриваются все элементы побега или их рудименты. Это все подземные структуры.
Менее распространены надземные видоизменения побега. Это, прежде всего, побеги суккулентов — растений, запасающих в паренхимных тканях воду. К таким растениям, кроме известных всем кактусов, относятся виды рода очиток, родиола (золотой корень), молодило и пр.
К надземным видоизменениям побега относятся стеблевые клубни (кольраби), кочан, образующийся у отдельных сортов капусты, уплощенные филлокладии иглицы, выводковые почки (мятлик луковичный, каланхоэ).
Жизненные формы
В ходе эволюции, у организмов сформировался определенный внешний облик — габитус. Особенности внешнего облика, сформировавшегося в результате адаптивного развития членов растения, получили название жизненных форм или биоморф. Таким образом, жизненная форма растения определяется морфологическим строением побега.
Различают три основные жизненные формы высших растений: деревья, кустарники, травы.
Деревья — это многолетние растения с покрытыми пробкой побегами, среди которых четко выделяется главный стебель — ствол. Высота деревьев от 40 см (карликовая береза) до 155 м (секвойя, эвкалипт).
Основные формы деревьев прямостоячие, но в пределах этой группы выделяются еще стланцы (кедровый стланник), у которых ствол стелится над почвой, и древесные лианы (виноград).
Кустарники — это многолетние растения, с покрытыми пробкой побегами без четко выраженного главного стебля. Осевые стебли кустарников называют стволиками. Высота кустарников колеблется от 0,5 до б м.
Низкорослые кустарники (от 5 до 60 см высоты) называют кустарничками.
Если у многолетнего многоосевого растения на зиму отмирает большая часть стволиков, то его называют полукустарником или полукустарничком, что зависит от его размеров. К полукустарникам относятся малина, многие виды полыни. Полукустарничком является чабрец.
Полукустарники с вьющимся стеблем называют полукустарниковыми лианами (паслен сладко-горький).
Травы подразделяются на многолетние, двулетние, однолетние.
Однолетние травы существуют один вегетационный период.
У двулетников в первый год образуется вегетативный побег, а на второй год происходит цветение и плодоношение, после чего растение погибает. У многолетних трав ежегодно погибает надземная часть, но сохраняется подземная. Такие растения переносят неблагоприятные условия (зима, сезон дождей в тропиках, засуха) в состоянии «корнеплодов», корнеклубней, корневищ, луковиц и т.д.
Редукция вегетативных органов у цветковых паразитов и полупаразитов
Растения-паразиты питаются готовыми органическими веществами, которые вырабатываются растениями-хозяевами. Из зародышевого корешка растений-паразитов образуется прокаул, который внедряется в корень растения-хозяина, откуда получает воду, минеральные и органические вещества. Листья растений-паразитов редуцированы до чешуек, стебель обычно мал, зато семенная продуктивность очень высокая. К растениям-паразитам относятся виды заразих, павилик, баланофоровых.
У растений-полупаразитов хорошо развита система побегов. Листья зеленые, обеспечивают образование органических веществ. От растения-хозяина растение-полупаразит получает воду с минеральными солями. Этот процесс обеспечивается прокаулом, т.е. у растений-полупаразитов редуцированным оказывается корень.
Морфология органов растения
В отличие от членов растения, которые выполняют комплекс функций, органы специализированы на выполнении одной конкретной функции.
Стробилы и цветки
В морфологии растений особое внимание уделяется таким органам, как стробилы и цветки.
Стробилы — собрание спорофиллов на оси, обособленной от вегетативной части растения. Стробилы характерны для растений из отделов плауновидные, хвощевидные, голосеменные. У плауновидных и хвощевидных их называют спороносными колосками. Развиваются они на концах побегов и состоят из оси и спорофиллов, несущих спорангии. У голосеменных стробилы — это мужские шишки и семенные чешуи женских шишек. Мужские шишки или мужские стробилы имеют ось и спорофиллы, которые несут спорангии. Женские стробилы или семенные чешуи женских шишек имеют ось и два спорофилла, сросшихся с осью, где каждый спорофилл несет по одному спорангию.
Цветок — это укороченный, неразветвленный побег с ограниченнным ростом, листья которого метаморфизированы в связи с адаптацией к размножению. Это особая морфологическая структура, присущая только покрытосеменным растениям.
Уникальность цветка по природе и функциям, существующие различные взгляды на его происхождение, привели к появлению противоречий в понимании общей структуры цветка, повлияли на нечеткость терминологии, применяемой при описании цветка.
В морфологической характеристике цветка мы исходили из его почечного происхождения. Цветок развивается из почки, а не из апекса почки, как утверждают некоторые морфологи. Ведь мы же не говорим, что побег развивается из апекса почки. Мы говорим, что он развивается из почки, а почка образуется в результате меристематической деятельности апекса.
В цветковой почке образуются бугорки, которые дают начало всем боковым элементам цветка. Осевые элементы цветка образуются из зачаточных узлов и междоузлий.
Исходя из вышеизложенного, полный цветок состоит из следующих частей: цветоножки, прицветников, прицветничка, цветоложа, околоцветника, подчашия, андроцея и гинецея.
Цветоножка — нижняя осевая часть цветка, бывает хорошо выражена или сильно укорочена, дифференцирована на три или на два междоузлия и иметь два или один узел. Может быть представлена одним междоузлием.
Прицветники — первые (обычно два) листочки, образующиеся из нижних листовых бугорков цветковой почки.
Прицветнички (обычно один) — листочки, образующиеся из второго снизу блока листовых бугорков.
Цветоложе — верхняя осевая часть цветка. Состоит из сильно сближенных узлов и очень коротких междоузлий. Количество узлов и междоузлий зависит от количества элементов и их расположения, которое может быть в виде ряда концентрических кругов, либо спиральным. Цветоложе может иметь различную форму и размеры.
Околоцветник — стерильная часть цветка. Он бывает двойным и простым. Двойной околоцветник дифференцирован на две части: чашечку и венчик, элементы которых — чашелистики и лепестки — резко отличаются друг от друга. Подчашие — листочки, расположенные на цветоложе под чашечкой, и отличающиеся от чашелистиков по форме и размерам. Простой околоцветник не дифференцирован на чашечку и венчик. Элементы простого околоцветника называются листочками.
Андроцей — совокупность тычинок.
Гинецей — совокупность плодолистиков, из которых образованы пестики, или пестик.
Цветок, у которого отсутствуют те или иные части, называется неполным.
Цветки, у которых нет цветоножки, называются сидячими.
Цветки без околоцветника называются беспокровными или голыми.
Цветки без андроцея называются женскими.
Цветки без гинецея называются мужскими. Цветки без гинецея и без андроцея называются стерильными.
Кроме того, морфологическое разнообразие цветков связано с количественными и качественными особенностями частей цветка.
Общие для всех цветков особенности связаны с характером расположения элементов околоцветника, тычинок, а иногда и плодолистиков на цветоложе, и с типом симметрии.
В зависимости от первого признака, цветки бывают циклическими, ациклическими (спиральными), гемициклическими (внизу кругами, вверху по спирали). Тип симметрии определяется для чашечки, венчика или для околоцветника в целом. В зависимости от этого признака цветки подразделяют на актиноморфные, или правильные, зигоморфные, или неправильные, и асимметричные.
Особый тип строения характерен для цветков злаков и осок. У злаков полный цветок состоит из оси, нижней цветковой чешуи, верхней цветковой чешуи, 2-3 лодикул, 3,6 тычинок и пестика, образованного из 2-3 сросшихся плодолистиков. У осок цветки состоят из оси, 6 или 3 чешуй, часто редуцированных до волосков или щетинок, трех тычинок и пестика из 2-3 плодолистиков. Если цветки однополые, то женский цветок заключен в мешочек — прицветник, сросшийся краями в колбовидное образование.
Структурная морфология пыльцы и семязачатка
Зрелая пыльца голосеменных имеет обычно бобовидную форму. У многих видов она снабжена двумя «воздушными мешками». Состоит из внешней толстой оболочки – экзины, и внутренней тонкой — интины. В полости пыльцевого зерна обычно расположены четыре клетки. Основной объем пыльцы занят вегетативной клеткой или, по современной терминологии, клеткой-трубкой. В основном веществе клетки-трубки находится генеративная клетка. Между интиной и клеткой-трубкой расположены две небольшие проталлиальные клетки.
Пыльца покрытосеменных имеет обычно округлую или эллиптическую форму. Внешняя толстая оболочка – экзина, часто снабжена различного вида шипиками или выростами другого характера, причем скульптура экзины является таксономическим признаком. Под экзиной находится внутренняя тонкая оболочка — интина. В полости зрелой пыльцы расположены две или три клетки. В первом случае весь объем пыльцы занят клеткой-трубкой, которую называют еще вегетативной клеткой, пыльцевой клеткой. В основном веществе клетки-трубки «плавает» генеративная клетка. Во втором случае в основном веществе цитоплазмы клетки-трубки расположены две клетки. Это половые клетки — спермии.
Все клетки пыльцы имеют гаплоидный набор хромосом.
Семязачаток голосеменных состоит из интегумента, нуцеллуса и женского гаметофита с погруженными в него двумя архегониями — женскими органами полового размножения. Эти три элемента как бы вложены один в другой. Интегумент — это наружный слой клеток семязачатка, его покров. На одном из полюсов интегумента расположено микропиле, под которым имеется небольшая полость — пыльцевая камера. Нуцеллус — ткань, находящаяся между интегументом и гаметофитом.
Гаметофит — это структура, продуцирующая гаметы. Часто вегетативную часть гаметофита у голосеменных называют эндоспермом, что является явной терминологической ошибкой. В архегониях образуются по одной яйцеклетке. Клетки интегумента и нуцеллуса содержат диплоидный набор хромосом, а клетки гаметофита — гаплоидный.
Семязачаток покрытосеменных состоит из интегумента, нуцеллуса и зародышевого мешка. Интегумент может быть однослойный или двуслойный, на верхушке семязачатка интегумент имеет микропиле (пыльцевход). Нуцеллус, расположенный между интегументом и зародышевым мешком, образует стенку зародышевого мешка. Зародышевый мешок занимает центральную часть семязачатка и состоит из семи клеток. Три клетки расположены в части зародышевого мешка, обращенной к микропиле. Это яйцевой аппарат. Яйцевой аппарат представлен яйцеклеткой и двумя клетками — синергидами. На противоположном полюсе зародышевого мешка сконцентрированы три клетки — антиподы. Основной объем зародышевого мешка представлен центральной двуядерной клеткой, которая плотно прилегает к антиподам — клеткам, образующим яйцевой аппарат, и к клеткам нуцеллуса. Ядра клеток зародышевого мешка гаплоидные.
Морфология гинецея
Гинецей — это совокупность плодолистиков. Плодолистики составляют основу пестика. Пестик обычно состоит из завязи, столбика и рыльца, реже пестик состоит из завязи и рыльца. Если в цветке есть много пестиков, то каждый из них образован одним плодолистиком, а совокупность этих пестиков называют апокарпным гинецеем. В этом случае завязь всегда бывает верхняя. Если в цветке пестик один, то гинецей может быть монокарпным, ценокарпным или псевдомонокарпным. Завязь у однопестичного (простого) гинецея может быть нижняя, верхняя или средняя.
Таксономическое значение имеют такие признаки гинецея, как количество столбиков, форма рыльца, наличие опушения, количество гнезд завязи и т.д.
Морфология соцветий
Соцветие — это группа цветков на специализированном побеге.
Морфология соцветий связана, прежде всего, с типом ветвления специализированного побега.
Отличаются соцветия количеством цветков, размерами, формой, наличием или отсутствием брактей и т.д.
В зависимости от ветвления различают соцветия ботриоидные (ботрические, неопределенные), которые характеризуются моноподиальным ветвлением, и цимоидные (цимозные, определенные), которые имеют ложнодихотомическое или симподиальное ветвление.
По степени сложности ветвления ботриоидные соцветия подразделяются на простые, сложные и агрегатные.
Основными составными частями простого соцветия являются ось и цветки. В пределах простых ботриоидных соцветий выделяют кистевидные, головковидные, колосовидные.
Цветки у кистевидных соцветий имеют цветоножки, а ось соцветия сравнительно тонкая. К этой группе соцветий относятся кисть, щиток, зонтик. Часто морфологи все кистевидные соцветия называют кистью, тогда говорят о кисти цилиндрической, кисти конической, кисти зонтиковидной, кисти головковидной, кисти щитковидной, кисти колосовидной и т.д.
Головковидные ботриоидные соцветия характерны для сложноцветных. Наиболее известным вариантом головки является корзинка. Тут характерный признак — разросшаяся ось и цветки на коротких цветоножках.
Колосовидные соцветия характеризуются наличием сидячих цветков.
Сложные ботриоидные соцветия имеют главную ось и парциальные соцветия. В качестве парциальных соцветий выступают простые ботриоидные соцветия. Отсюда и название сложных соцветий: сложная кисть, сложный щиток, сложный зонтик. Особым типом сложного ботриоидного соцветия является антела. Это соцветие, по-видимому, является производным сложного щитка, у которого междоузлия нижних щитков сильно увеличены, а количество цветков сокращено до двух.
Ботриоидные соцветия, где ветвление по типу кисти сочетается с парциальными соцветиями — зонтик и корзинка, называют агрегатными. Это метелка из зонтиков, метелка из корзинок, щиток из корзинок, кисть из корзинок.
Особым видом ботриоидных соцветий являются соцветия злаков. Парциальное соцветие злаков — колосок, который развивается в пазухе кроющего листа — нижней колосковой чешуи и состоит из оси, брактей и 1-2-многих цветков. Колоски собраны в сложные метельчатые, колосовидные, головкообразные, зонтикообразные общие соцветия.
Цимоидные соцветия подразделяются на два типа: цимоиды и тирсы.
Цимоиды — это простые и сложные цимоидные соцветия. Простые цимоиды — это монохазий и дихазий. Монохазий имеет два цветка, дихазий — три. Первый образуется за счет симподиального ветвления, второй за счет ложнодихотомического.
Сложные цимоиды представляют собой ветвящиеся симподиально простые цимоиды (завиток, извилина, плейохазий, плейохазий двойной) или ветвящиеся ложнодихотомические цимоиды (двойной дихазий, тройной дихазий и т.д.).
Особый тип цимоидного соцветия — циатий встречается у молочайных. Такое соцветие образовано одним пестичным и пятью тычиночными цветками.
Тирсы — это цимоидные соцветия, образованные простыми и сложными циатиями, расположенными на осях второго порядка в основании тирса, и на оси первого порядка на вершине тирса.
Морфология семян
Семена у семенных растений образуются из семязачатка.
Семена голосеменных имеют плотную семенную кожуру, иногда снабженную крыловидным выростом. Под семенной кожурой расположена пленчатая структура — остаток нуцеллуса. Остальной объем семени занят разросшимся телом гаметофита, трансформированным в питательную ткань, и зародышем. Зародыш расположен в специальной камере. Состоит зародыш из корешка, стебелька, семядолей и почечки. С питательной тканью зародыш связан подвеском, который отходит от зародышевого корешка.
Семена покрытосеменных сильно различаются по весу, размерам, окраске, рисунку поверхности.
Морфологические особенности позволяют выделить четыре типа семян.
1. Семена с эндоспермом. Эндосперм — это питательная ткань, которая имеется только у покрытосеменных. Развивается из триплоидного ядра, образовавшегося в процессе двойного оплодотворения, когда вначале сливаются ядра центральной клетки, а с образовавшимcя диплоидным ядром сливается ядро спермия. Такие семена состоят из семенной кожуры, эндосперма и зародыша.
2. Семена с эндоспермом и периспермом. Перисперм — это питательная ткань, образовавшаяся в результате специализации нуцеллуса. В этом случае семена состоят из семенной кожуры, перисперма, эндосперма и зародыша.
3. Семена с периспермом. Такие семена состоят из семенной кожуры, перисперма и зародыша.
4. Семена без эндосперма и перисперма. У таких семян есть семенная кожура и зародыш.
Морфология плодов
Плод — это зрелый цветок. Из цветка с одним пестиком образуются простые плоды. Из цветка с несколькими пестиками образуются, так называемые, сборные или сложные плоды. Парциальной частью сборного плода является плодик, который образуется из одного пестика.
Плоды и плодики состоят из околоплодника и семян, или из околоплодника и одного семени. Околоплодник, в свою очередь, дифференцирован на три части: экзокарп — внешняя часть; мезокарп – средняя часть, и эндокарп — внутренняя часть.
Морфологическое разнообразие плодов связано с их размерами, окраской, формой, с наличием различных приспособлений к распространению, с консистенцией частей околоплодника, с количеством гнезд, семян и т.д.
Сравнительно-морфологические исследования используются в классификации плодов. Общепринятыми являются морфологическая классификация, которая используется до сего времени в фармакогнозии, и морфогенетическая классификация.
Морфогенетическая классификация приведена в базовом учебнике. Тут будет рассмотрена морфологическая классификация.
Простые и сборные плоды подразделяются на сухие и мясистые, или сочные.
Главные виды простых мясистых плодов — костянка, ягода, яблоко, тыквина, гесперидий. У костянки эндокарп каменистый, мезокарп мясистый, экзокарп пленчатый. Это, обычно, односеменные плоды. Двусеменная костянка у кизила. У костянки кокосового ореха — мезокарп волокнистый. Все остальные виды мясистых плодов — многосеменные. У ягоды — мясистые мезокарп и эндокарп. У яблока — мясистый мезокарп, жесткопленчатый эндокарп, кожистый экзокарп. У тыквины — мясистый мезокарп и деревянистый экзокарп. У гесперидия — железистый экзокарп, губчатый мезокарп и сочный эндокарп.
Сухие плоды бывают односеменными, дву- и многосеменными.
К сухим простым плодам относятся семянки — одногнездные плоды со свободным семенем; зерновки — плоды, у которых семя срастается с околоплодником; орехи — плоды с каменистой стенкой; схизокарпии — плоды, распадающиеся при созревании на две или более односеменных частей.
Листовки — многосеменные плоды, вскрывающиеся по одному шву; бобы — многосеменные плоды, вскрывающиеся по двум швам; стручки — плоды двугнездные, вскрывающиеся двумя створками; коробочки – многосеменные плоды, вскрывающиеся трещинами, зубцами, дырочками.
Сборные сухие плоды — это многолистовка — многосеменные плодики, вскрываются по одному шву; многоорешек — плодики односеменные, не вскрываются; цинародий — плод шиповников; фрага — плод земляники. В принципе, плоды шиповников и земляники — это многоорешки с разросшимся мясистым цветоложем.
К сборным мясистым плодам относится многокостянка и сочная многолистовка (лимонник китайский).
Морфология соплодий
Соплодие — несколько плодов, образовавшихся из отдельных цветков соцветия, сросшихся между собой. Это соплодие шелковицы, соплодие инжира, соплодие свеклы, соплодие ананаса.
Морфологическая эволюция покрытосеменных растений
Основные направления структурной эволюции отражены в следующих эволюционно-морфологических рядах.
Жизненные формы
Вечнозеленые растения — листопадные растения
Деревья – кустарники — травы многолетние — травы однолетние.
Корневые системы
Система главного корня — смешанная корневая система — система придаточных корней
Стебли
Ветвистые — простые
Прямостоячие — восходящие (приподнимающиеся) — стелющиеся (лежачие)
Ветвление моноподиальное — ветвление ложнодихотомическое — ветвление симподиальное
Лист и листорасположение
Листья простые — листья сложные
Листья сложные — листья вторично простые
Листья сидячие — листья черешковые
Листья с прилистниками — листья без прилистников
Листовая пластинка цельная — листовая пластинка пальчато-расчлененная — листовая пластинка перисто-расчлененная
Листья простые с пальчато-расчлененной пластинкой — листья пальчатосложные — листья тройчатосложные — листья перистосложные
Листья простые с пальчато-расчлененной пластинкой — листья с перисто-расчлененной пластинкой — листья перистосложные — листья тройчатосложные
Листья простые с пальчато-расчлененной пластинкой — листья пальчатосложные — листья перистосложные — листья тройчатосложные
Листья простые с цельной пластинкой — листья тройчато-расчлененные — листья тройчатосложные — листья перистосложные.
Листья с ланцетной листовой пластинкой — … — листья с округлой листовой пластинкой
Листья с пальчатым жилкованием — листья с перистым жилкованием.
Листья с пальчатым жилкованием — листья с дуговым жилкованием — листья с параллельным жилкованием.
Листорасположение очередное — листорасположение супротивное — листорасположение мутовчатое.
Цветок и соцветие
Цветки одиночные — цветки в ботриоидных соцветиях — цветки в цимоидных соцветиях
Цветки обоеполые — цветки однополые — цветки стерильные
Цветки актиноморфные — цветки зигоморфные — цветки асимметричные
Спиральное расположение частей цветка — спиральноциклическое расположение частей цветка — циклическое расположение частей цветка
Число элементов частей цветка большое и неопределенное — число элементов частей цветка небольшое и фиксированное
Околоцветник двойной — околоцветник простой — околоцветника нет
Элементы частей цветка свободные — элементы частей цветка сросшиеся
Пестики (совокупность плодолистиков, гинецей) на ножках — пестики сидячие
Пестиков в цветке много — пестик в цветке один
Завязь верхняя — завязь средняя — завязь нижняя
Семя
Семена с двумя семядолями — семена с одной семядолей
Семена с эндоспермом — семена без эндосперма
Семена с эндоспермом — семена с эндоспермом и периспермом — семена с периспермом — семена без эндосперма и перисперма
Плоды
Плоды сборные — плоды простые
Эволюционно-морфологические ряды используются в качестве примеров примитивности или специализации структур при создании систем цветковых растений.
Данные сравнительно-морфологических исследований позволяют создать фенетические системы. Создание же генеалогических систем требует использования данных большинства разделов ботаники: анатомии, биохимии и т.д. Учитывая, что основным методом эволюционной систематики все-таки остается морфолого-географический, пока рассмотрим основы этого раздела ботаники.
Часть ботаники
Фитоморфология — это изучение физической формы и внешнего строения растений. растения. Это обычно считается отличным от анатомии растений, которая представляет собой изучение внутренней структуры растений, особенно на микроскопическом уровне. Морфология растений полезна для визуальной идентификации растений.
 Соцветия, появляющиеся из защитного покрова
Соцветия, появляющиеся из защитного покрова
Содержание
- 1 Область применения
- 2 Сравнительная наука
- 2.1 Гомология
- 2.2 Конвергенция
- 3 Вегетативные и репродуктивные характеристики
- 3.1 Использование при идентификации
- 3.2 Смена поколений
- 3.3 Пигментация у растений
- 4 Морфология в развитии
- 4.1 Рост
- 4.2 Морфологическая изменчивость
- 4.3 Эволюция морфологии растений
- 4.3.1 Позиционные эффекты
- 4.3.2 Воздействие окружающей среды
- 4.3.2.1 Температура
- 4.3.3 Молодость
- 5 Современная морфология растений
- 6 См. Также
- 7 Ссылки
- 8 Внешние ссылки
Объем
 Asclepias syriaca, демонстрирующая сложную морфологию цветов.
Asclepias syriaca, демонстрирующая сложную морфологию цветов.  Изучение структуры ветви дерева Pinus sylvestris
Изучение структуры ветви дерева Pinus sylvestris
Морфология растений «представляет собой исследование развития, формы и строения растений и, как следствие, попытки интерпретировать их на основе сходства плана и происхождения ». Морфология растений состоит из четырех основных областей, каждая из которых совпадает с другой областью биологических наук.
Прежде всего, морфология сравнительная, что означает, что морфолог изучает структуры многих растений одного и того же или разных видов, затем проводит сравнения и формулирует идеи о сходстве. Когда считается, что структуры у разных видов существуют и развиваются в результате общих унаследованных генетических путей, эти структуры называют гомологичными. Например, листья сосны, дуба и капусты выглядят очень по-разному, но имеют общую структуру и расположение частей.. О гомологии листьев легко сделать. Морфолог растений идет дальше и обнаруживает, что колючки кактуса также имеют такую же базовую структуру и развитие, что и листья других растений, и поэтому колючки кактуса также гомологичны листьям. Этот аспект морфологии растений пересекается с изучением эволюции и палеоботаники.
. Во-вторых, морфология растений учитывает обе вегетативные (соматические ) структуры. растений, а также репродуктивных структур. Вегетативные структуры сосудистых растений включают исследование системы побегов, состоящей из стеблей и листьев, а также системы корней.. Репродуктивные структуры более разнообразны и обычно характерны для определенной группы растений, например цветы и семена, папоротник сори и мох. капсулы. Детальное изучение репродуктивных структур растений привело к открытию чередования поколений, обнаруженных у всех растений и большинства водорослей. Эта область морфологии растений пересекается с изучением биоразнообразия и систематики растений.
В-третьих, морфология растений изучает структуру растений в различных масштабах. В самых маленьких масштабах: ультраструктура, общие структурные особенности клеток, видимые только с помощью электронного микроскопа, и цитология, исследование клеток с помощью оптической микроскопии. В этом масштабе морфология растений пересекается с анатомией растений как областью исследования. В самом крупном масштабе проводится изучение растения привычки роста, общей архитектуры растения. Схема ветвления дерева будет варьироваться от вида к виду, как и внешний вид растения в виде дерева, травы или травы..
В-четвертых, морфология растений исследует образец развития, процесс, в результате которого структуры возникают и созревают по мере роста растения. В то время как животные производят все части тела, которые они когда-либо будут иметь с раннего возраста, растения постоянно производят новые ткани и структуры на протяжении всей своей жизни. У живого растения всегда есть зародышевые ткани. На то, как созревают новые структуры по мере их создания, может влиять момент в жизни растения, когда они начинают развиваться, а также среда, воздействию которой подвергаются структуры. Морфолог изучает этот процесс, причины и его результат. Эта область морфологии растений пересекается с физиологией растений и экологией.
Сравнительная наука
Морфолог растений сравнивает структуры многих разных растений одного и того же или разных видов. Проведение таких сравнений между похожими структурами на разных заводах решает вопрос, почему эти структуры похожи. Вполне вероятно, что подобные основные причины генетики, физиологии или реакции на окружающую среду привели к этому внешнему сходству. Результат научного исследования этих причин может привести к одному из двух выводов о лежащей в основе биологии:
- Гомология — структура схожа у двух видов из-за общего происхождения и общей генетики.
- Конвергенция — структура похожа у двух видов из-за независимой адаптации к общим воздействиям окружающей среды.
Понимание того, какие характеристики и структуры принадлежат каждому типу, является важной частью понимания эволюции растений . Биолог-эволюционист полагается на морфолога растений для интерпретации структур и, в свою очередь, обеспечивает филогении взаимоотношений растений, которые могут привести к новым морфологическим открытиям.
Гомология
Когда считается, что структуры у разных видов существуют и развиваются в результате общих наследственных генетических путей, эти структуры называют гомологичными. Например, листья сосны, дуба и капусты выглядят по-разному, но имеют общие основные структуры и расположение частей. Сделать вывод о гомологии листьев несложно. Морфолог растений идет дальше и обнаруживает, что колючки кактуса также имеют ту же основную структуру и развитие, что и листья других растений, и поэтому колючки кактуса также гомологичны листьям.
Конвергенция
Когда считается, что структуры у разных видов существуют и развиваются в результате общих адаптивных реакций на давление окружающей среды, эти структуры называют конвергентными. Например, листья Bryopsis plumosa и стебли Asparagus setaceus имеют одинаковый перистый ветвящийся вид, хотя один из них является водорослью, а другой — цветковым растением. Сходство в общей структуре происходит независимо в результате конвергенции. Формы роста многих кактусов и видов молочай очень похожи, хотя они принадлежат к очень далеким семействам. Сходство является результатом общих решений проблемы выживания в жаркой и сухой среде.
Вегетативные и репродуктивные характеристики
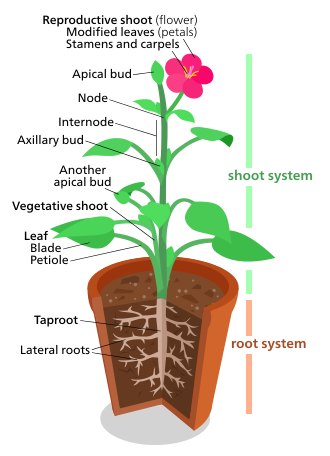 Диаграмма, представляющая «типичный» эвдикот.
Диаграмма, представляющая «типичный» эвдикот.
Морфология растений касается как вегетативных структур растений, так и репродуктивных структур.
вегетативные (соматические ) структуры сосудистых растений включают две основные системы органов: (1) систему побегов, состоящий из стеблей и листьев, и (2) корневой системы . Эти две системы являются общими почти для всех сосудистых растений и представляют собой объединяющую тему для изучения морфологии растений.
Напротив, репродуктивные структуры разнообразны и обычно специфичны для конкретной группы растений. Такие структуры, как цветы и плоды встречаются только у покрытосеменных ; сори встречаются только в папоротниках ; и семенные шишки встречаются только у хвойных деревьев и других голосеменных растений. Поэтому репродуктивные признаки считаются более полезными для классификации растений, чем вегетативные признаки.
Использование в идентификации
Биологи растений используют морфологические признаки растений, которые можно сравнивать, измерять, подсчитывать и описывать, для оценки различий или сходств в таксонах растений и используют эти признаки для идентификации и классификации растений и описания.
Когда символы используются в описаниях или для идентификации, они называются диагностическими или ключевыми символами, которые могут быть как качественными, так и количественными.
- Количественные признаки — это морфологические признаки, которые можно подсчитать или измерить, например, у вида растений лепестки цветков шириной 10–12 мм.
- Качественные признаки — это морфологические признаки, такие как форма листьев, цвет или опушение цветка.
Оба типа символов могут быть очень полезны для идентификации растений.
Смена поколений
Детальное изучение репродуктивных структур у растений привело к открытию смены поколений, обнаруженной у всех растений и большинства водорослей, немецким ботаником Вильгельмом Хофмайстером. Это открытие — одно из самых важных, сделанных во всей морфологии растений, поскольку оно обеспечивает общую основу для понимания жизненного цикла всех растений.
Пигментация растений
Основная функция пигментов в растениях — фотосинтез, при котором используется зеленый пигмент хлорофилл вместе с несколькими красными и желтыми пигментами. которые помогают улавливать как можно больше световой энергии. Пигменты также являются важным фактором в привлечении насекомых к цветам, чтобы стимулировать опыление.
Растительные пигменты включают множество различных типов молекул, включая порфирины, каротиноиды, антоцианы и беталаины. Все биологические пигменты избирательно поглощают одни длины волн света, в то время как отражают другие. Поглощенный свет может быть использован растением для стимуляции химических реакций, в то время как отраженные длины волн света определяют цвет, который пигмент будет казаться глазу.
Морфология в развитии
Растение Развитие — это процесс, посредством которого структуры возникают и созревают по мере роста растения. Это предмет изучения анатомии растений и физиологии растений, а также морфологии растений.
Процесс развития у растений фундаментально отличается от того, что наблюдается у позвоночных животных. Когда животное эмбрион начинает развиваться, оно очень рано производит все части тела, которые когда-либо будут у него в жизни. Когда животное рождается (или вылупляется из яйца), у него есть все части тела, и с этого момента оно будет только расти и становиться более зрелым. Напротив, растения на протяжении всей своей жизни постоянно производят новые ткани и структуры из меристем, расположенных на концах органов или между зрелыми тканями. Таким образом, у живого растения всегда есть зародышевые ткани.
Свойства организации, наблюдаемые в растении, — это возникающие свойства, которые больше, чем сумма отдельных частей. «Сборка этих тканей и функций в единый многоклеточный организм дает не только характеристики отдельных частей и процессов, но и совершенно новый набор характеристик, которые нельзя было бы предсказать на основе исследования отдельных частей». Другими словами, знания всего о молекулах в растении недостаточно, чтобы предсказать характеристики клеток; а знание всех свойств клеток не позволяет предсказать все свойства структуры растения.
Рост
A сосудистого растения начинается с одноклеточной зиготы, образованной оплодотворением яйцеклетки сперматозоидом. С этого момента он начинает делиться с образованием зародыша растения в процессе эмбриогенеза . Когда это происходит, полученные клетки организуются так, что один конец становится первым корнем, а другой конец — верхушкой побега. В семенных растениях из зародыша разовьется один или несколько «семенных листьев» (семядоли ). К концу эмбриогенеза у молодого растения будут все части, необходимые для начала его жизни.
Как только зародыш прорастает из своего семени или родительского растения, он начинает производить дополнительные органы (листья, стебли и корни) в процессе органогенеза . Новые корни вырастают из корневых меристем, расположенных на кончике корня, а новые стебли и листья растут из меристем побега, расположенных на кончике побега. Ветвление происходит, когда небольшие скопления клеток, оставленные меристемой и еще не подвергшиеся клеточной дифференцировке с образованием специализированной ткани, начинают расти как верхушка нового корня или побега. Рост любой такой меристемы на кончике корня или побега называется первичным ростом и приводит к удлинению этого корня или побега. Вторичный рост приводит к расширению корня или побега за счет делений клеток в камбии.
Помимо роста за счет деления клеток, растение может вырасти через удлинение ячейки . Это происходит, когда отдельные клетки или группы клеток становятся длиннее. Не все растительные клетки вырастают до одинаковой длины. Когда клетки на одной стороне ствола растут дольше и быстрее, чем клетки на другой стороне, в результате ствол будет изгибаться в сторону более медленно растущих клеток. Этот направленный рост может происходить через реакцию растения на определенный стимул, такой как свет (фототропизм ), гравитация (гравитропизм ), вода (гидротропизм ), и физический контакт (тигмотропизм ).
Рост и развитие растений опосредуются специфическими гормонами растений и регуляторами роста растений (PGR) (Ross et al. 1983). Уровни эндогенных гормонов зависят от возраста растений, морозостойкости, состояния покоя и других метаболических состояний; фотопериод, засуха, температура и другие внешние условия окружающей среды; и экзогенные источники ГРР, например, внесенные извне и ризосферного происхождения.
Морфологическая изменчивость
Растения проявляют естественные вариации по своей форме и структуре. В то время как все организмы различаются от человека к человеку, растения демонстрируют дополнительный тип изменений. В пределах одного человека повторяются части, которые могут отличаться по форме и структуре от других подобных частей. Это изменение наиболее легко увидеть на листьях растения, хотя другие органы, такие как стебли и цветы, могут иметь аналогичные изменения. У этой вариации есть три основные причины: позиционные эффекты, влияние окружающей среды и молодость.
Эволюция морфологии растений
Факторы транскрипции и сети регуляции транскрипции играют ключевую роль в морфогенезе растений и их эволюции. Во время посадки растений появилось много новых семейств факторов транскрипции, которые преимущественно связаны с сетями многоклеточного развития, воспроизводства и развития органов, что способствует более сложному морфогенезу наземных растений.
Позиционные эффекты
 Вариации в листьях из гигантской амброзии, иллюстрирующей позиционные эффекты. Лопастные листья исходят из основания растения, а недоливные листья — из верхушки растения.
Вариации в листьях из гигантской амброзии, иллюстрирующей позиционные эффекты. Лопастные листья исходят из основания растения, а недоливные листья — из верхушки растения.
Хотя растения производят множество копий одного и того же органа в течение своей жизни, не все копии конкретного органа будут идентичными. Есть различия между частями зрелого растения, обусловленные относительным положением органа. Например, на новой ветке листья могут иметь одинаковый узор вдоль ветки. Форма листьев, образующихся у основания ветки, будет отличаться от листьев, образующихся на кончике растения, и это различие сохраняется от ветки к ветке на данном растении и в данном виде. Это различие сохраняется после того, как листья на обоих концах ветки созрели, и не является результатом того, что одни листья моложе других.
Воздействие окружающей среды
На то, как созревают новые структуры по мере их создания, может влиять момент в жизни растений, когда они начинают развиваться, а также среда, в которой они конструкции выставлены. Это можно увидеть в водных растениях и надводных растениях.
Температура
Температура оказывает множество эффектов на растения в зависимости от множества факторов, включая размер и состояние растений. растение, а также температуру и продолжительность воздействия. Чем меньше и суккулентнее растение, тем больше подвержено повреждению или гибели от слишком высоких или слишком низких температур. Температура влияет на скорость биохимических и физиологических процессов, скорость обычно (в определенных пределах) увеличивается с температурой. Однако соотношение Вант-Гоффа для мономолекулярных реакций (которое гласит, что скорость реакции удваивается или утроится при повышении температуры на 10 ° C) не строго соблюдается для биологических процессов, особенно при низких и высоких температурах.
Когда вода замерзает в растениях, последствия для растений во многом зависят от того, происходит ли замерзание внутриклеточно (внутри клеток) или вне клеток в межклеточном (внеклеточном) пространстве. Внутриклеточное замораживание обычно приводит к гибели клетки вне зависимости от морозостойкости растения и его тканей. Внутриклеточное замораживание редко встречается в природе, но умеренные скорости снижения температуры, например, от 1 ° C до 6 ° C / час, вызывают образование межклеточного льда, и этот «внеклеточный лед» может быть или не быть смертельным, в зависимости от выносливости. ткани.
При отрицательных температурах сначала замерзает вода в межклеточных пространствах тканей растений, хотя вода может оставаться незамерзшей до тех пор, пока температура не упадет ниже 7 ° C. После первоначального межклеточного образования льда клетки сжимаются, поскольку вода теряется в сегрегированном льду. Клетки подвергаются сублимационной сушке, причем обезвоживание является основной причиной обморожения.
Было показано, что скорость охлаждения влияет на морозостойкость тканей, но фактическая скорость замораживания будет зависеть не только от скорости охлаждения, но также от степени переохлаждения и свойств ткани. Сакаи (1979a) продемонстрировал сегрегацию льда в зачатках побегов белой и черной елей Аляски при медленном охлаждении до 30–40 ° C. Эти обезвоженные замораживанием почки выжили при погружении в жидкий азот при медленном нагревании. Аналогичным образом отреагировали и цветочные зачатки. Внеорганное промерзание в зачатках объясняет способность наиболее выносливых из северных хвойных деревьев переживать зимы в регионах, где температура воздуха часто опускается до -50 ° C и ниже. Зимостойкость зимних почек таких хвойных деревьев повышается за счет небольшого размера почек, эволюции более быстрого перемещения воды и способности переносить интенсивное обезвоживание замораживанием. У бореальных видов Picea и Pinus морозостойкость однолетних проростков не уступает зрелым растениям при аналогичных состояниях покоя.
Молодость
 Молодость в проростке бука европейского. Существует заметная разница в форме между первыми темно-зелеными «семенными листьями» и более светлой второй парой листьев.
Молодость в проростке бука европейского. Существует заметная разница в форме между первыми темно-зелеными «семенными листьями» и более светлой второй парой листьев.
Органы и ткани, вырабатываемые молодым растением, например саженец, часто бывают отличается от тех, которые производятся тем же самым заводом, когда он старше. Это явление известно как ювенильность или гетеробластика. Например, молодые деревья будут давать более длинные и тонкие ветви, которые будут расти вверх больше, чем ветви, которые они будут производить как полностью выросшее дерево. Кроме того, листья, образовавшиеся на раннем этапе роста, обычно крупнее, тоньше и неправильной формы, чем листья взрослого растения. Образцы молодых растений могут настолько отличаться от взрослых растений того же вида, что яйцекладущие насекомые не распознают растение как пищу для своего потомства. Различия проявляются в укоренении и цветении, а также на одном и том же зрелом дереве. Молодые черенки, взятые у основания дерева, образуют корни гораздо легче, чем черенки, исходящие из средней и верхней кроны. Цветение у основания дерева отсутствует или менее обильно, чем цветение на более высоких ветвях, особенно когда молодое дерево впервые достигает возраста цветения.
Переход от ранних форм роста к поздним обозначается как «изменение вегетативной фазы ‘, но есть некоторые разногласия по поводу терминологии.
Современная морфология растений
Рольф Саттлер пересмотрел фундаментальные концепции сравнительной морфологии, такие как концепция гомологии. Он подчеркнул, что гомология должна также включать частичную гомологию и количественную гомологию. Это приводит к континуальной морфологии, которая демонстрирует континуум между морфологическими категориями корня, побега, стебля (каулом), листа (филлом) и волос (трихома). Как лучше всего описываются промежуточные звенья между категориями, обсуждали Bruce K. Kirchoff et al. Недавнее исследование, проведенное Stalk Institute, извлекло координаты, соответствующие основанию и листьям каждого растения в трехмерном пространстве. Когда растения на графике были размещены в соответствии с их фактическим расстоянием перемещения питательных веществ и общей длиной ветвей, растения почти идеально попадали на кривую Парето. «Это означает, что способ выращивания растений в своей архитектуре также оптимизирует очень распространенный компромисс при проектировании сети. В зависимости от окружающей среды и вида, завод выбирает различные способы достижения компромиссов для этих конкретных условий окружающей среды».
В честь Агнес Арбер, автора теории частичных побегов листа, Рутисхаузер и Ислер назвали континуальный подход нечеткой арберианской морфологией (FAM). «Нечеткое» относится к нечеткой логике, «Арбериан» — к Агнес Арбер. Рутисхаузер и Ислер подчеркнули, что этот подход подтверждается не только многими морфологическими данными, но и данными молекулярной генетики. Более свежие данные молекулярной генетики предоставляют дополнительную поддержку морфологии континуума. Джеймс (2009) пришел к выводу, что «в настоящее время широко признано, что… радиальность [характеристика большинства стеблей] и дорсивентральность [характеристика листьев] — всего лишь крайности непрерывного спектра. Фактически, это просто синхронизация гена KNOX. выражение !. » Эккард и Баум (2010) пришли к выводу, что «в настоящее время общепринято, что сложные листья выражают свойства как листа, так и побега».
Морфология процесса описывает и анализирует динамический континуум формы растения. Согласно этому подходу, структуры действительно не имеют процесса (ов), они являются процессом (процессами). Таким образом, дихотомия структура / процесс преодолевается за счет «расширения нашего понятия« структура »с тем, чтобы включить и признать, что в живом организме это не просто вопрос о пространственной структуре с «активностью» как чем-то над или против нее, но что конкретный организм является пространственно-временной структурой и что эта пространственно-временная структура является самой деятельностью ».
Для Жен, Барабе и Лакруа классическая морфология (то есть основная морфология, основанная на концепции качественной гомологии, подразумевающей взаимоисключающие категории) и морфология континуума являются подклассами более всеобъемлющей морфологии процесса (динамической морфологии).
Классический мо Рфология, морфология континуума и морфология процесса имеют большое значение для эволюции растений, особенно в области эволюционной биологии растений (растение evo-DevO), которая пытается объединить морфологию растений и молекулярную генетику растений. В подробном тематическом исследовании необычных морфологий Рутисхаузер (2016) проиллюстрировал и обсудил различные темы эволюции растений, такие как нечеткость (непрерывность) морфологических концепций, отсутствие взаимно однозначного соответствия между структурными категориями и экспрессией генов., понятие морфопространства, адаптивная ценность особенностей bauplan по сравнению с патио ludens, физиологическая адаптация, обнадеживающие монстры и сальтационная эволюция, значение и пределы устойчивости развития и т. д.
Нравится нам это или нет, морфологические исследования находится под влиянием философских допущений, таких как либо / или логика, нечеткая логика, дуализм структуры / процесса или его трансцендентность. А эмпирические данные могут повлиять на философские предположения. Таким образом, существует взаимодействие между философией и эмпирическими открытиями. Эти взаимодействия являются предметом того, что было названо философией морфологии растений.
См. Также
- Глоссарий морфологии растений
- Анатомия растений
- Идентификация растений
- Физиология растений
- Эволюционная биология развития растений
- Таксономия
Ссылки
Внешние ссылки
- Ботанический визуальный глоссарий
- Морфология растений: фундаментальные принципы
Фитоморфология это исследование физическая форма и внешнее строение из растения.[1] Обычно это считается отличным от анатомия растений,[1] который является изучением внутреннего структура растений, особенно на микроскопическом уровне.[2] Морфология растений полезна для визуальной идентификации растений. Недавние исследования в молекулярная биология начал исследовать молекулярные процессы, участвующие в определении сохранения и разнообразия морфологии растений. В этих исследованиях транскриптом образцы сохранения были обнаружены, чтобы отметить важные онтогенетический переходы во время завод жизненный цикл, который может привести к эволюционным ограничениям, ограничивающим диверсификацию[3].
Объем
Морфология растений «представляет собой исследование развития, формы и структуры растений и, как следствие, попытку интерпретировать их на основе сходства плана и происхождения».[4] Морфология растений состоит из четырех основных областей, каждая из которых частично совпадает с другой областью исследований. Биологические науки.
Прежде всего, морфология сравнительныйЭто означает, что морфолог изучает структуры многих разных растений одного или разных видов, затем проводит сравнения и формулирует идеи о сходстве. Когда считается, что структуры у разных видов существуют и развиваются в результате общих, унаследованных генетический пути, эти структуры называются гомологичный. Например, уходит из сосна, дуб, и капуста все они выглядят по-разному, но имеют общую структуру и расположение частей. О гомологии листьев легко сделать. Морфолог растений идет дальше и обнаруживает, что шипы из кактус также имеют ту же базовую структуру и развитие, что и листья у других растений, поэтому колючки кактуса также гомологичны листьям. Этот аспект морфологии растений частично совпадает с изучением растений. эволюция и палеоботаника.
Во-вторых, морфология растений учитывает как вегетативный (соматический) конструкции растений, а также репродуктивный конструкции. Вегетативные структуры сосудистые растения включает изучение системы побегов, состоящей из стебли и уходит, так же хорошо как корень система. Репродуктивные структуры более разнообразны и обычно характерны для определенной группы растений, таких как цветы и семена, папоротник сори, и мох капсулы. Детальное изучение репродуктивных структур растений привело к открытию смена поколений содержится во всех растениях и большинстве водоросли. Эта область морфологии растений частично совпадает с изучением биоразнообразие и систематика растений.
В-третьих, морфология растений изучает структуру растений в различных масштабах. В самых маленьких масштабах ультраструктура, общие структурные особенности клеток видны только с помощью электронный микроскоп, и цитология, исследование клеток с помощью оптическая микроскопия. В этом масштабе морфология растений пересекается с анатомия растений как область исследования. В самом крупном масштабе — это изучение растений. привычка к росту, общая архитектура завода. Схема ветвления в дерево будет варьироваться от вида к виду, как и внешний вид растения как дерево, трава, или трава.
В-четвертых, морфология растений изучает характер развитие, процесс, посредством которого структуры возникают и созревают по мере роста растения. В то время как животные производят все части тела, которые они когда-либо будут иметь с раннего возраста, растения постоянно производят новые ткани и структуры на протяжении всей своей жизни. У живого растения всегда есть зародышевые ткани. На то, как созревают новые структуры по мере их создания, может влиять момент в жизни растения, когда они начинают развиваться, а также среда, которой подвергаются эти структуры. Морфолог изучает этот процесс, причины и его результат. Эта область морфологии растений пересекается с физиология растений и экология.
Сравнительная наука
Морфолог растений сравнивает структуры многих растений одного или разных видов. Проведение таких сравнений между подобными структурами на разных заводах решает вопрос Зачем конструкции похожи. Вполне вероятно, что подобные основные причины генетики, физиологии или реакции на окружающую среду привели к этому внешнему сходству. Результат научного исследования этих причин может привести к одному из двух открытий лежащей в основе биологии:
- Гомология — структура схожа у двух видов из-за общего происхождения и общей генетики.
- Конвергенция — структура похожа у двух видов из-за независимой адаптации к общим воздействиям окружающей среды.
Понимание того, какие характеристики и структуры принадлежат каждому типу, является важной частью понимания растений. эволюция. Биолог-эволюционист полагается на морфолога растений для интерпретации структур и, в свою очередь, дает филогении взаимоотношений растений, которые могут привести к новым морфологическим открытиям.
Гомология
Когда считается, что структуры у разных видов существуют и развиваются в результате общих наследственных генетических путей, эти структуры называют гомологичный. Например, листья сосны, дуба и капусты выглядят по-разному, но имеют общие основные структуры и расположение частей. О гомологии листьев легко сделать. Морфолог растений идет дальше и обнаруживает, что колючки кактуса также имеют ту же основную структуру и развитие, что и листья других растений, и поэтому колючки кактуса также гомологичны листьям.
Конвергенция
Когда считается, что структуры у разных видов существуют и развиваются в результате общих адаптивных реакций на давление окружающей среды, эти структуры называют сходящийся. Например, листья Bryopsis plumosa и основы Спаржа щетинистая оба имеют одинаковые перистые ветвления, хотя одна из них — водоросль, а другая — цветущее растение. Сходство в общей структуре происходит независимо в результате конвергенции. Форма роста многих кактусы и виды Молочай очень похожи, хотя и принадлежат к очень далеким семьям. Сходство является результатом общих решений проблемы выживания в жаркой и сухой среде.
Вегетативные и репродуктивные характеристики
Диаграмма, представляющая «типичный» эвдикот.
Морфология растений касается как вегетативных структур растений, так и репродуктивных структур.
В вегетативный (соматический) структуры сосудистые растения включают две основные системы органов: (1) система стрельбы, состоящий из стебли и уходит, и (2) a корневая система. Эти две системы являются общими почти для всех сосудистых растений и представляют собой объединяющую тему для изучения морфологии растений.
Напротив, репродуктивный структуры разнообразны и обычно характерны для определенной группы растений. Такие структуры как цветы и фрукты находятся только в покрытосеменные; сори находятся только в папоротники; и шишки находятся только в хвойные породы и другие голосеменные. Поэтому репродуктивные признаки считаются более полезными для классификации растений, чем вегетативные признаки.
Использование в идентификации
Биологи растений используют морфологические признаки растений, которые можно сравнивать, измерять, подсчитывать и описывать, чтобы оценить различия или сходства в таксонах растений и использовать эти признаки для идентификации, классификации и описания растений.
Когда символы используются в описаниях или для идентификации, они называются диагностический или ключевые персонажи которые могут быть как качественными, так и количественными.
- Количественные признаки — это морфологические признаки, которые можно подсчитать или измерить, например, у вида растений лепестки цветов шириной 10–12 мм.
- Качественные признаки — это морфологические признаки, такие как форма листьев, цвет цветка или опушение.
Оба типа символов могут быть очень полезны для идентификации растений.
Смена поколений
Детальное изучение репродуктивных структур растений привело к открытию смена поколений, встречается во всех растениях и большинстве водоросли, немецкий ботаник Вильгельм Хофмайстер. Это открытие — одно из самых важных, сделанных во всей морфологии растений, поскольку оно обеспечивает общую основу для понимания жизненного цикла всех растений.
Пигментация у растений
Основная функция пигментов в растениях — фотосинтез, который использует зеленый пигмент хлорофилл вместе с несколькими красными и желтыми пигментами, которые помогают улавливать как можно больше световой энергии. Пигменты также являются важным фактором привлечения насекомые к цветам, чтобы ободрить опыление.
Растительные пигменты включают множество различных видов молекул, в том числе: порфирины, каротиноиды, антоцианы и беталаины. Все биологические пигменты избирательно поглощают определенные длины волн свет в то время как отражающий другие. Поглощаемый свет может использоваться растением для стимуляции химических реакций, а длина отраженной волны света определяет цвет, который пигмент будет казаться глазом.
Морфология в развитии
Завод развитие это процесс, посредством которого структуры возникают и созревают по мере роста растения. Это предмет изучения в анатомия растений и физиология растений а также морфология растений.
Процесс развития растений принципиально отличается от того, что наблюдается у растений. позвоночное животное животные. Когда животное эмбрион начинает развиваться, он очень рано производит все части тела, которые когда-либо будут у него в жизни. Когда животное рождается (или вылупляется из яйца), у него есть все части тела, и с этого момента оно будет только расти и становиться более зрелым. Напротив, растения на протяжении всей своей жизни постоянно производят новые ткани и структуры из меристемы[5] расположены на концах органов или между зрелыми тканями. Таким образом, у живого растения всегда есть зародышевые ткани.
Свойства организации, наблюдаемые в растении: эмерджентные свойства которые больше, чем сумма отдельных частей. «Сборка этих тканей и функций в единый многоклеточный организм дает не только характеристики отдельных частей и процессов, но и совершенно новый набор характеристик, которые нельзя было бы предсказать на основе исследования отдельных частей».[6] Другими словами, знания всего о молекулах в растении недостаточно, чтобы предсказать характеристики клеток; а знание всех свойств клеток не позволяет предсказать все свойства структуры растения.
Рост
А сосудистое растение начинается с одноклеточной зигота, образована оплодотворение яйцеклетки с помощью сперматозоида. С этого момента он начинает делиться, образуя растение. эмбрион через процесс эмбриогенез. Когда это произойдет, полученные клетки будут организованы так, что один конец станет первым корнем, а другой конец — верхушкой побега. В семя растения, у эмбриона разовьется один или несколько «семенных листков» (семядоли ). К концу эмбриогенеза у молодого растения будут все части, необходимые для начала его жизни.
Однажды эмбрион прорастает из своего семени или родительского растения он начинает производить дополнительные органы (листья, стебли и корни) в процессе органогенез. Новые корни растут из корня меристемы расположен на кончике корня, и из побега вырастают новые стебли и листья меристемы находится на кончике побега.[7] Ветвление происходит, когда небольшие скопления клеток, оставленные меристемой и еще не подвергшиеся клеточная дифференциация чтобы сформировать специализированную ткань, начинают расти как верхушка нового корня или побега. Рост из любой такой меристемы на кончике корня или побега называется первичный рост и приводит к удлинению этого корня или побега. Вторичный рост приводит к расширению корня или побега из делений клеток в камбий.[8]
Помимо роста на ячейка деление, растение может прорасти удлинение ячейки. Это происходит, когда отдельные клетки или группы клеток становятся длиннее. Не все растительные клетки вырастают до одинаковой длины. Когда клетки на одной стороне ствола растут дольше и быстрее, чем клетки на другой стороне, в результате ствол будет изгибаться в сторону более медленно растущих клеток. Этот направленный рост может происходить через реакцию растения на определенный стимул, такой как свет (фототропизм ), сила тяжести (гравитропизм ), воды, (гидротропизм ) и физический контакт (тигмотропизм ).
Рост и развитие растений опосредуются специфическими гормоны растений и регуляторы роста растений (ГРР) (Росс и др., 1983).[9] Уровни эндогенных гормонов зависят от возраста растений, морозостойкости, состояния покоя и других метаболических состояний; фотопериод, засуха, температура и другие внешние условия окружающей среды; и экзогенные источники ГРР, например, применяемые извне и ризосферного происхождения.
Морфологическая вариация
Растения демонстрируют естественные вариации по форме и строению. В то время как все организмы различаются от человека к человеку, растения демонстрируют дополнительный тип изменений. В пределах одного человека повторяются части, которые могут отличаться по форме и структуре от других подобных частей. Это изменение наиболее легко увидеть на листьях растения, хотя другие органы, такие как стебли и цветы, могут иметь аналогичные изменения. У этой вариации есть три основные причины: позиционные эффекты, влияние окружающей среды и молодость.
Эволюция морфологии растений
Факторы транскрипции и сети регуляции транскрипции играют ключевую роль в морфогенезе растений и их эволюции. Во время посадки растений появилось много новых семейств факторов транскрипции, которые преимущественно связаны с сетями многоклеточного развития, репродукции и развития органов, внося вклад в более сложный морфогенез наземных растений.[10]
Позиционные эффекты
Вариация листьев гигантской амброзии, иллюстрирующая позиционные эффекты. Лопастные листья исходят из основания растения, а недоливные — из верхушки растения.
Хотя растения производят множество копий одного и того же органа в течение своей жизни, не все копии одного и того же органа будут идентичными. Существуют различия между частями зрелого растения, обусловленные относительным положением органа. Например, на новой ветке листья могут иметь одинаковый узор вдоль ветки. Форма листьев, образующихся у основания ветки, будет отличаться от листьев, образующихся на верхушке растения, и эта разница сохраняется от ветки к ветке на данном растении и в данном виде. Это различие сохраняется после того, как листья на обоих концах ветки созрели, и не является результатом того, что одни листья моложе других.
Экологические последствия
На то, как созревают новые структуры по мере их создания, может влиять момент в жизни растений, когда они начинают развиваться, а также среда, которой подвергаются эти структуры. Это можно увидеть в водные растения и надводные растения.
Температура
Температура оказывает множество воздействий на растения в зависимости от множества факторов, включая размер и состояние растения, а также температуру и продолжительность воздействия. Чем меньше и больше сочные растения, тем выше подверженность повреждению или смерти от слишком высоких или слишком низких температур. Температура влияет на скорость биохимических и физиологических процессов, скорость обычно (в определенных пределах) увеличивается с температурой. Однако соотношение Вант-Гоффа для мономолекулярных реакций (которое гласит, что скорость реакции удваивается или утроится при повышении температуры на 10 ° C) не строго соблюдается для биологических процессов, особенно при низких и высоких температурах.
Последствия замерзания воды в растениях во многом зависят от того, происходит ли замерзание внутриклеточно (внутри клеток) или вне клеток в межклеточном (внеклеточном) пространстве.[11] Внутриклеточное замораживание обычно приводит к гибели клетки вне зависимости от морозостойкости растения и его тканей.[12] Внутриклеточное замораживание в природе встречается редко, но умеренные скорости снижения температуры, например, от 1 ° C до 6 ° C / час, вызывают образование межклеточного льда, и этот «внеклеточный лед»[13] может быть или не быть летальным, в зависимости от прочности ткани.
При отрицательных температурах сначала замерзает вода в межклеточных пространствах тканей растений, хотя вода может оставаться незамерзшей до тех пор, пока температура не упадет ниже 7 ° C.[11] После первоначального межклеточного образования льда клетки сжимаются, поскольку вода теряется в сегрегированном льду. Клетки подвергаются сублимационной сушке, причем обезвоживание является основной причиной обморожения.
Показано, что скорость охлаждения влияет на морозостойкость тканей,[14] но фактическая скорость замораживания будет зависеть не только от скорости охлаждения, но также от степени переохлаждения и свойств ткани.[15] Сакаи (1979a)[14] продемонстрировал сегрегацию льда в зачатках побегов белой и черной елей Аляски при медленном охлаждении до 30–40 ° C. Эти замороженные и обезвоженные почки пережили погружение в жидкий азот при медленном согревании. Аналогичным образом отреагировали и цветочные зачатки. Внеорганное промерзание в зачатках объясняет способность наиболее выносливых из северных хвойных деревьев переживать зимы в регионах, где температура воздуха часто опускается до -50 ° C и ниже.[13] Зимостойкость почек таких хвойные породы усиливается за счет небольшого размера почек, более быстрого перемещения воды и способности переносить интенсивное обезвоживание замораживанием. У бореальных видов Picea и Pinus, морозостойкость однолетних сеянцев на уровне зрелых растений,[16] учитывая подобные состояния покоя.
Юность
Молодость в проростке Европейский бук. Существует заметная разница в форме между первыми темно-зелеными «семенными листьями» и более светлой второй парой листьев.
Органы и ткани, производимые молодым растением, например, саженец, часто отличаются от тех, которые производятся тем же растением, когда они стареют. Это явление известно как юность или гетеробластика. Например, молодые деревья будут давать более длинные и тонкие ветви, которые будут расти вверх больше, чем ветви, которые они будут производить как полностью выросшее дерево. Кроме того, листья, образовавшиеся на раннем этапе роста, обычно крупнее, тоньше и неправильной формы, чем листья взрослого растения. Образцы молодых растений могут настолько отличаться от взрослых растений того же вида, что яйцекладущие насекомые не распознают растение как пищу для своего потомства. Различия видны в укоренении и цветении, а также на одном и том же зрелом дереве. Молодые черенки, взятые у основания дерева, образуют корни гораздо легче, чем черенки, исходящие из средней и верхней кроны. Цветение у основания дерева отсутствует или менее обильно, чем цветение на более высоких ветвях, особенно когда молодое дерево впервые достигает возраста цветения.[17]
Переход от ранних форм роста к поздним обозначается как «изменение вегетативной фазы ‘, но есть некоторые разногласия по поводу терминологии.[18]
Современная морфология растений
Рольф Саттлер пересмотрел фундаментальные концепции сравнительной морфологии, такие как концепция гомология. Он подчеркнул, что гомология должна также включать частичную гомологию и количественную гомологию.[19][20] Это приводит к континуальной морфологии, которая демонстрирует континуум между морфологическими категориями корня, побега, стебля (каулом), листа (филлом) и волос (трихома). Как лучше всего описываются промежуточные звенья между категориями, обсуждали Bruce K. Kirchoff et al.[21] Недавнее исследование, проведенное Stalk Institute, извлекло координаты, соответствующие основанию и листьям каждого растения в трехмерном пространстве. Когда растения на графике были размещены в соответствии с их фактическим расстоянием перемещения питательных веществ и общей длиной ветвей, растения почти идеально попадали на кривую Парето. «Это означает, что способ выращивания растений в своей архитектуре также оптимизирует очень распространенный компромисс при проектировании сети. В зависимости от окружающей среды и вида, завод выбирает различные способы достижения компромиссов для этих конкретных условий окружающей среды». [22]
В честь Агнес Арбер, автора теория частичного побега листа, Рутисхаузер и Ислер назвали континуальный подход нечеткой арберовской морфологией (FAM). «Нечеткий» относится к нечеткая логика, «Арбериан» в Агнес Арбер. Рутисхаузер и Ислер подчеркнули, что этот подход подтверждается не только многими морфологическими данными, но и данными молекулярной генетики.[23] Более свежие данные молекулярной генетики предоставляют дополнительную поддержку морфологии континуума. Джеймс (2009) пришел к выводу, что «в настоящее время широко признано, что … радиальность [характеристика большинства стеблей] и дорсивентральность [характеристика листьев] — всего лишь крайности непрерывного спектра. Фактически, это просто синхронизация гена KNOX. выражение !. «[24] Эккардт и Баум (2010) пришли к выводу, что «сейчас общепринято, что сложные листья выражают свойства как листьев, так и побегов».[25]
Морфология процесса описывает и анализирует динамический континуум формы растений. Согласно этому подходу конструкции не имеют процесс (ы), они находятся процесс (ы).[26][27][28] Таким образом, дихотомия структура / процесс преодолевается «расширением нашего понятия« структура », чтобы включить и признать, что в живом организме это не просто вопрос пространственной структуры с« активностью »как чем-то сверх или против. это, но что конкретный организм — это пространственно-временный структура и что эта пространственно-временная структура и есть сама деятельность «.[29]
Для Жен, Барабе и Лакруа классическая морфология (то есть основная морфология, основанная на концепции качественной гомологии, подразумевающей взаимоисключающие категории) и морфология континуума являются подклассами более всеобъемлющей морфологии процесса (динамической морфологии).[30]
Классическая морфология, морфология континуума и морфология процесса очень важны для эволюции растений, особенно в области эволюционной биологии растений (evo-DevO), которая пытается объединить морфологию растений и молекулярную генетику растений.[31] В подробном тематическом исследовании необычных морфологий Рутисхаузер (2016) проиллюстрировал и обсудил различные темы эволюции растений, такие как нечеткость (непрерывность) морфологических концепций, отсутствие взаимно однозначного соответствия между структурными категориями и экспрессией генов. , понятие морфопространства, адаптивная ценность особенностей bauplan по сравнению с патио ludens, физиологические адаптации, обнадеживающие монстры и сальтационная эволюция, значение и пределы устойчивости развития и т. д.[32]
Нравится нам это или нет, но морфологические исследования находятся под влиянием философских допущений, таких как либо / или логика, нечеткая логика, дуализм структуры / процесса или его трансцендентность. А эмпирические данные могут повлиять на философские предположения. Таким образом, существует взаимодействие между философией и эмпирическими открытиями. Эти взаимодействия являются предметом того, что было названо философией морфологии растений.[33]
Смотрите также
- Словарь морфологии растений
- Анатомия растений
- Идентификация растений
- Физиология растений
- Эволюционная биология развития растений
- Таксономия
использованная литература
- ^ а б Рэйвен П. Х., Р. Ф. Эверт и С. Э. Эйххорн. Биология растений, 7-е изд., Стр. 9. (Нью-Йорк: У. Х. Фриман, 2005). ISBN 0-7167-1007-2.
- ^ Эверт, Рэй Франклин и Исав, Кэтрин (2006) Анатомия растения Исава: меристемы, клетки и ткани растительного тела — их структура, функции и развитие Уайли, Хобокен, Нью-Джерси, страница xv, ISBN 0-471-73843-3
- ^ Дрост, Хайк-Георг; Белльштедт, Джулия; Ó’Maoiléidigh, Diarmuid S .; Сильва, Андерсон Т .; Габель, Александр; Вайнхольдт, Клаус; Райан, Патрик Т .; Dekkers, Bas J.W .; Бенцинк, Леони; Hilhorst, Henk W.M .; Лигтеринк, Уилко; Веллмер, Фрэнк; Гроссе, Иво; Квинт, Марсель (23 февраля 2016 г.). «Постэмбриональные модели песочных часов отмечают онтогенетические переходы в развитии растений». Молекулярная биология и эволюция. 33 (5): 1158–1163. Дои:10.1093 / molbev / msw039. PMID 26912813.
- ^ Гарольд К. Болд, К. Дж. Алексопулос и Т. Делеворяс. Морфология растений и грибов, 5-е изд., Стр. 3. (Нью-Йорк: Харпер-Коллинз, 1987). ISBN 0-06-040839-1.
- ^ Bäurle, I; Ло, Т. (2003). «Апикальные меристемы: фонтан молодости растений». BioEssays. 25 (10): 961–70. Дои:10.1002 / bies.10341. PMID 14505363. Обзор.
- ^ Леопольд, А.С. Рост и развитие растений, стр. 183. (Нью-Йорк: McGraw-Hill, 1964).
- ^ Бренд, U; Хобе, М; Саймон, Р. (2001). «Функциональные домены в меристемах побегов растений». BioEssays. 23 (2): 134–41. Дои:10.1002 / 1521-1878 (200102) 23: 2 <134 :: AID-BIES1020> 3.0.CO; 2-3. PMID 11169586. Обзор.
- ^ Барлоу, П. (2005). «Узорчатое определение клеток в растительной ткани: вторичная флоэма деревьев». BioEssays. 27 (5): 533–41. Дои:10.1002 / bies.20214. PMID 15832381.
- ^ Ross, S.D .; Pharis, R.P .; Биндер, У. Д. 1983. Регуляторы роста и хвойные деревья: их физиология и потенциальное использование в лесном хозяйстве. п. 35–78 в Никелл, Л. (Ред.), Химические вещества, регулирующие рост растений. Vol. 2, CRC Press, Бока-Ратон, Флорида.
- ^ Jin JP; и другие. (Июль 2015 г.). «Карта регуляторов транскрипции Arabidopsis выявляет отличительные функциональные и эволюционные особенности новых факторов транскрипции». Молекулярная биология и эволюция. 32 (7): 1767–1773. Дои:10.1093 / molbev / msv058. ЧВК 4476157. PMID 25750178.
- ^ а б Glerum, C. 1985. Морозостойкость саженцев хвойных пород: принципы и применение. п. 107–123 в Дурья, М. (Ред.). Ход работы: Оценка качества рассады: принципы, процедуры и прогностические возможности основных тестов. Workshop, октябрь 1984 г., Университет штата Орегон, For. Res. Lab., Corvallis OR.
- ^ Lyons, J.M .; Raison, J.K .; Степонкус, П. 1979. Растительная мембрана в ответ на низкую температуру: обзор. п. 1–24 в Lyons, J.M .; Graham, D .; Raison, J.K. (Ред.). Низкотемпературный стресс у сельскохозяйственных культур. Academic Press, Нью-Йорк, штат Нью-Йорк.
- ^ а б Sakai, A .; Ларчер, В. (ред.) 1987. Морозостойкость растений. Springer-Verlag.
- ^ а б Сакаи, A. 1979a. Механизм предотвращения промерзания примордиальных побегов почек хвойных. Physiol растительных клеток. 20: 1381–1390.
- ^ Левитт, Дж. 1980. Реакция растений на стрессы окружающей среды. Том 1. Переохлаждение, замерзание и высокотемпературные напряжения, 2-е изд. Academic Press, Нью-Йорк, штат Нью-Йорк. 497 с.
- ^ Sakai, A .; Окада, С. 1971. Морозостойкость хвойных пород. Silvae Genet. 20 (3): 91–97.
- ^ Майкл А. Дирр; Чарльз Хойзер-младший (2006). «2». Справочное руководство по размножению древесных растений (Второе изд.). Varsity Press Inc., стр. 26, 28, 29. ISBN 0942375092.
- ^ Джонс, Синтия С. (1999-11-01). «Очерк молодости, фазового изменения и гетеробластики семенных растений». Международный журнал наук о растениях. 160 (S6): –105 – S111. Дои:10.1086/314215. ISSN 1058-5893. PMID 10572025.
- ^ Саттлер Р. (1984). «Гомология — непрекращающийся вызов». Систематическая ботаника. 9 (4): 382–394. Дои:10.2307/2418787. JSTOR 2418787.
- ^ Саттлер Р., 1994, Гомология, гомеоз и морфология процессов у растений. В: Б.К. Холл (ред.) Гомология: иерархическая основа сравнительной морфологии. Нью-Йорк: Academic Press, стр. 423-475.
- ^ Кирхгоф, Б. К.; Pfeifer, E; Рутисхаузер, Р. (2008). «Онтология структуры растений: как мы должны маркировать структуры растений с сомнительной или смешанной идентичностью?». Zootaxa. 1950: 103–122. Дои:10.11646 / zootaxa.1950.1.10.
- ^ Конн, Адам; Педмале, Уллас; Чори, Джоанн (2017). «Лазерное сканирование с высоким разрешением показывает, что архитектура предприятия отражает универсальные принципы проектирования сети». Сотовые системы. 5 (1): 103–122. Дои:10.1016 / j.cels.2017.06.017. PMID 28750198.
- ^ Rutishauser, R .; Ислер, Б. (2001). «Генетика развития и морфологическая эволюция цветковых растений, особенно пузырчатых (Utricularia): нечеткая арберовская морфология дополняет классическую морфологию» (PDF). Анналы ботаники. 88 (6): 1173–1202. Дои:10.1006 / anbo.2001.1498.
- ^ Джеймс, П. Дж. (2009). «‘Дерево и лист: Другой ракурс ». Линней. 25: 13–19.
- ^ Eckardt, NA; Баум, Д. (2010). «Загадка Podostemad: эволюция необычной морфологии у Podostemaceae». Растительная клетка. 22 (7): 2131–2140. Дои:10.1105 / tpc.110.220711. ЧВК 2929115. PMID 20647343.
- ^ Саттлер, Р. (1992). «Морфология процессов: структурная динамика в развитии и эволюции». Канадский журнал ботаники. 70 (4): 708–714. Дои:10.1139 / b92-091.
- ^ Вергара-Силва, Ф. (2003). «Растения и концептуальная артикуляция эволюционной биологии развития». Биология и философия. 18 (2): 261–264. Дои:10.1023 / А: 1023936102602. S2CID 81013686.
- ^ Саттлер, Р. 2019. Структурные и динамические подходы к развитию и эволюции формы растений. В: Fusco, G. (ed) Перспективы эволюционной биологии и биологии развития. Очерки для Алессандро Минелли. Глава 6, с. 57-70 [1]
- ^ Вудгер, Дж. 1967 г. Биологические принципы. Лондон: Рутледж и Кегоан Пол (переиздан с новым Введением).
- ^ Jeune, B; Barabé, D; Лакруа, С. (2006). «Классическая и динамическая морфология: к синтезу через пространство форм». Acta Biotheoretica. 54 (4): 277–293. Дои:10.1007 / s10441-007-9007-8. PMID 17486414. S2CID 25928998.
- ^ Минелли, А. 2018. Эволюционная биология развития растений. Эволюционируемость фенотипа. Нью-Йорк: Издательство Кембриджского университета.
- ^ Рутисхаузер, Р. (2016). «Эволюция необычных морфологий у Lentibulariaceae (пузырчатые и родственные им) и Podostemonaceae (речные сорняки): графическое изображение на стыке биологии развития и морфологической диверсификации». Анналы ботаники. 117 (5): 811–832. Дои:10.1093 / aob / mcv172. ЧВК 4845801. PMID 26589968.
- ^ Саттлер, Р. (2018). «Философия морфологии растений». Elemente der Naturwissenschaft. 108: 55–79.(расширенную версию этой статьи см. [2] )
внешние ссылки
- Ботанический визуальный глоссарий
- Морфология растений: основные принципы
Integrated Views in Plant Breeding
Grazia M. Borrelli, … Luigi Cattivelli, in Crop Physiology, 2009
4.2.3 Morphological and developmental modifications
Plant morphology and architecture and the transcriptional networks controlling plant development have been targeted in yield improvement studies. Examples concern the modification of root growth, by affecting cell expansion, and of leaf and stem architecture, by acting on gibberellin’s and brassinosteroid’s signalling pathways (Van Camp, 2005). The identification of the genes responsible for plant dwarfing, related to plant response to gibberellin, or affecting the formation of tillers and lateral branches carrying inflorescences has opened promising ways for yield improvement (Takeda et al., 2003; Sakamoto and Matsuoka, 2004). The alteration of phytochrome phyA and phyB gene expression also represents a powerful strategy to alter plant morphology. By means of phytochrome photoreceptors, plants sense the reduction in the ratio of red (R) to far-red (FR) light when they are shaded by their neighbours and respond by increasing their stem extension, reducing branching, accelerating leaf senescence and reducing allocation of resources to yield organs (Smith, 2000; Libenson et al., 2002). Transgenic overexpression of oat phyA in tobacco showed an enhanced allocation of assimilates to leaves with a consequent increase in fresh leaf harvest index, particularly when plants were grown at high density in the field (Robson et al., 1996). Positive effects on leaf morphology, photosynthetic performance and tuber yield were found in transgenic potatoes overexpressing Arabidopsis phyB (Thiele et al., 1999; Schittenhelm et al., 2004) that were confirmed in field studies at high plant population density (Boccalandro et al., 2003). The enhanced phyB expression not only altered the ability of plants to respond to light signals, but also modified the light environment itself, resulting in higher maximum photosynthesis in leaves of all strata of the canopy, largely due to increased leaf stomatal conductance, and a yield increase due to the increased tuber number.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780123744319000141
Moth bean
Ramavtar Sharma, … Man Mohan Sundria, in The Beans and the Peas, 2021
4.3 Biology and crop phenology
The plant morphology differs considerably in different genotypes of moth bean (Fig. 4.1). With short internodes, the partially angular stem grows upto 30 cm. Dense-packed branched moth bean plant can reach a span of up to 150 cm. Both stem and branches are striated with long, spreading and deflexed hairs. The leaflets of trifoliated leaves are variable in size and shape and more or less deeply lobed. Lobes are oblong, acute or acuminate, more or less hairy when young but become glabrescent with age. The inflorescence is raceme on axillary 5–6 cm long peduncle with yellowish hairs. Racemes are capitated with setaceous and ciliated tips protruding beyond the buds (Brink and Jansen, 2006). Bisexual flowers are papilionaceous and the calyx is campanulated and 4–5 mm long. Being a member of subgenera Ceratotropis (Asiatic Vigna), moth bean flowers always exhibits different shades of yellow color (Baudoin and Marechal, 1988). The yellowish corolla is 5–5.5 mm long. The keel is sickle-shaped. Ten stamens are arranged in nine united and one free fashion. With incurved style, the ovary is sessile and superior. Fruits are cylindrical pods (2.4 × 0.5 cm) having curved beak covered with short stiff hairs. Pod color is brown or pale gray when mature and has 5–8 seed. Rectangular to cylindrical seeds, occasionally reniform shaped, 4.5 × 3 mm are whitish-green, yellow to brown often mottled with black. The linear whitish hilum is nonprotruding. Epigeal germination is rule in moth bean (Majumdar, 2011).
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780128214503000044
THE WORLD OF FOOD GRAINS
J. Wyatt, in Encyclopedia of Food Grains (Second Edition), 2016
Introduction
The definition of plant morphology is the physical appearance of a plant. It can be applied to any species and involves a detailed study of vegetative and reproductive characters in order to form a profile of a plant, which can be used to make general comparisons of plant species displaying a similar structure or detailed comparisons within a species to identify varieties (cultivars). Identifying varieties and species of plants or grains by morphological means is still the main method used in cereal breeding and varietal identification and provides the basis for the definitive description of a variety.
Plants within a species share a similar genetic base, and thus, it is possible to group them into a taxonomic family. ‘Cereals’ is a generic term used to encompass the major grain-producing species whose products are some of the world’s most important staple foods and have been since man started growing them for food some 12 000 years ago. A cereal variety is a population of plants that can be identified as being clearly distinguishable from another population by stable inherited morphological characters. Wheat (Triticum aestivum), rice (Oryza sativa), and maize (Zea mays) account for over half of the cereals consumed worldwide. Cereals sit in the family Poaceae (synonymous with Gramineae). While cereals differ in detail in the way the characters of each species are expressed, they share a general commonality in growth habit and flowering parts. By understanding the morphology of each species, it is possible to use it to identify the varieties (cultivars) that exist in each classification. Recognition of the importance attached to identifying varieties is highlighted by the existence of UPOV (The International Union for the Protection of New Plant Varieties). The test guidelines put together by the 71 members of UPOV provide useful information of the major characters used in establishing the morphological identification of the different varieties of cereal species.
The following section begins with a description of the main morphological characters common to all the cereal species and is followed by further sections specific to each species. Illustrations and photographs are used to help in the identification of the characters described.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780123944375000097
Trichoderma-plant-pathogen interactions for benefit of agriculture and environment
Narendra Kumar, S.M. Paul Khurana, in Biocontrol Agents and Secondary Metabolites, 2021
3.3 Effect on plant physiology, effect on yield and quality of produce
The Trichoderma spp. have the power to changing the plant physiology, viz., net photosynthetic rate, stomatal conductance, internal CO2 concentration, transpiration, nutrient uptake, and water use efficiency. Thus, nutrient uptake enhances physiological processes in plants that lead to good growth (Doni et al., 2014). Micronutrients such as Nitrogen and Magnesium are the components of chlorophyll and enzymes of m-RNA synthesis and gene regulation engaged in photosynthesis. Trichoderma harzianum significantly increased the ability of rice plants to tolerate drought stress and increase in rice plant’s water-holding capacity (Shukla et al., 2012). An approximately threefold increase in the net photosynthetic rate and stomatal conductance and twofold increase in water use efficiency in Trichoderma-treated rice plants as compared to NPK-treated plants has been observed (Doni et al., 2014). The increased photosynthetic rates with less transpiration in Trichoderma treats projects efficiency of high water use (Doni et al., 2014). The high photosynthetic potential in Trichoderma-treated plants with less CO2 concentration can be correlated as an activity of carboxylation through CO2 fixation for glucose production at the time of carbohydrate metabolism, which is very active (Thakur et al., 2010). The prolonged photosynthetic activity supporting delayed senescence in rice is due to activity of Trichoderma spp. (Mishra and Salokhe, 2011). Trichoderma releases cellulases which degrade cellulose and enhance the organic matter and nutrients in the rhizosphere. Jiang et al. (2011) reported that increase of plant physiological processes such as solubilization and minerals chelation may enhance nutrient availability that can upgrade plant metabolism (Harman et al., 2004).
3.3.1 Effect on yield and quality of produce
Trichoderma effect on seed germination, plant morphology, and physiology leads to a better field stand and it also accelerates the vegetative and reproductive growth of plants. It enhances the number of branches, spikes, flowers, and fruits per plant. In many cases, the average weight of individual fruit is also comparatively higher. Higher yield by the application of Trichoderma species in mustard, wheat, corn, tuberose, sugarcane, tomato, okra, etc. has been found (Haque et al., 2012; El-Katatny and Idres, 2014; Naznin et al., 2015; Idowu et al., 2016).
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B978012822919400003X
Inter- and Intraspecific Morphological Variation in Bumblebee Species, and Competition in Flower Utilization
Tamiji Inoue, Makoto Kato, in Effects of Resource Distribution on Animal–Plant Interactions, 1992
C Plant Characters That Relate to Pollination
From the viewpoint of plants, plant morphology is generally thought to affect fertilization success, by increasing pollinator visitation and by decreasing improper pollen transfer (Rathcke, 1983). Visitation intensity of insects is basically determined by the attractiveness of plants (Chapter 5), or the net energetic intake from the plant per unit time (Pleasants, 1980). Rare plants produce much more nectar per flower than do abundant plants to increase their attractiveness (Heinrich, 1976a, b, c; Pleasants, 1980). Flower morphology adds a level of specificity to attractiveness by excluding noneffective pollinators, or robbers. Long flowers protect nectar from short-tongued bees and, as exemplified by Rabdosia longituba, characters other than flower depth may also determine the accessibility of flowers to bees. Such exclusion mechanisms have been thought not to be present in short and dish-shaped flowers, but this is not necessarily true (Kakutani et al., 1989). Short flowers of the Japanese orchid, Platanthera nipponica, for example, that are pollinated by short-tongued noctuids, exclude long-tongued sphingids by having twisted spurs, because only straight spurs can be probed by the long tongues of hovering sphingids (Inoue, 1983; another example in Impatiens in Kato et al., 1991). Neither type can escape from piercing by primary nectar robbers (Inouye, 1980; Roubik, 1989, 1990, see Chapter 11).
Intraspecific size variation is also found in flowers, and different-sized flowers within a single plant species attract different-sized bees. Larger flowers of cow vetch, Vicia cracca, were visited by larger workers of Bombus vagans, and vice versa (Morse, 1978). Assortive mating among similar size classes was found in this plant. Galen and Newport (1987) investigated optimal flower design in sky pilot, Polemonium viscosum. Larger flowers were advantageous because they increased the number of visits by pollinators (higher reward in large flowers), but suffered from a lower frequency of precise pollen transfer. Optimization of flower size results from stabilizing selection in this plant, which balances the opposing forces. By changing spur lengths of orchids experimentally, Inoue (1987) found that shortened spurs decreased both pollinia removal rate (paternal success) and fertilization rate (maternal success). There were no significant deleterious effects of elongating spurs and, in this case, longer flowers would be favored. Our study did not deal with intraspecific size variation of plants. We suggest that simultaneous studies of size variation of both plants and bees would prove fruitful.
The positions at which pollen or pollinia are attached to pollinator bodies are determined by flower morphology. Sharing of a single pollinator by multiple plant species becomes possible if pollen deposition sites vary among the plants (Waser, 1983; Macior, 1982; Feinsinger, 1987; Kato et al., 1991). Sharing pollinators in this way is especially important for rare plants (Rathcke, 1983) to receive the service of trapliners (Janzen, 1971).
However, inter- and intraspecific morphological variation among plants does not always imply specialization of pollinator species specific to individual plant species or morphs (Chapter 11). There was considerable interspecific variation, for example, in the flower morphology of North American Pedicularis, but each species was visited by several common bumblebee species (Macior, 1982, but he did not measure bee morphology). Even specific morphological characters that seem specialized for a single pollinator are frequently pillaged by robbers, parasites, and others (Roubik, 1989; Chapter 11).
Color and odor are other floral characters that advertise flowers to pollinators (Keven, 1978; Macior, 1978). Intraspecific color morphs can promote assortive matings (Brown and Clegg, 1984), while intraspecific odor morphs, each of which is pollinated differentially by bumblebees and flies, are also known (Galen et al., 1987). The classification of pollinator syndromes based on color and odor is found in classical works of pollination ecology.
Flowering phenology has been studied frequently in relation to competition for pollination (Rathcke, 1983). Both staggering of flowering periods among plant members in the same guild that share a single pollinator (Pleasants, 1980; Heinrich, 1976b; Yumoto, 1986, 1987), and random associations (Rathcke, 1984; Tepedino and Stanton, 1981, 1982) are reported from different communities. Such wide variation among the results, which depends on the communities studied and the researchers’ prejudice, demands more careful examination of the patterns. For this purpose, methodologies for the statistical testing of data with null models have been developed by various authors (Pleasants, 1980; Rathcke, 1984). Pleasants (1990) examined by several methods the flowering times of plant species that were hypothetically staggered, and found that his original analysis of the plant–bee communities in the Rocky Mountains (Pleasants, 1980), by mean pairwise overlap, was statistically correct. But in Pleasants (1980), the problem is not in the statistical analysis, but in the rather arbitrary procedure used to assign plants to specific guilds. Boundaries between guilds are generally not distinct, but gradual, even in Pleasants (1980) data. We need a statistically rational standard to discriminate among guilds (Jaksic and Medel, 1990). Such a classification of plants according to pollinator syndromes tends, in many cases, to exaggerate specialization (Waser, 1983).
In the present study, we used cluster analysis for the classification of plants by visitation, but we could not detect distinct guilds of plants visited by four out of five bumblebee species. As shown below, in combinations of two or three plant species, overlap of flowering periods decreases seed set (see Chapter 5 for a further discussion of apparent competition among plants for pollinators generated by differences in nectar production rates). At the community level, however, staggering of flowering periods among plant species appears rare, and is incomplete if present. In addition, the phylogeny of plants (Kochmer and Handel, 1986) and habitat conditions (Macior, 1983) limit the free allocation of flowering periods by plants that is a basic assumption of null models. Simultaneous analyses of both morphology and phenology have been suggested (Murray et al., 1987).
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780080918815500165
Plant Macrofossil Introduction☆
H.H. Birks, in Reference Module in Earth Systems and Environmental Sciences, 2017
How Will Plant Macrofossil Analysis Develop in the Future?
Macrofossil analysis requires good botanical knowledge of plant morphology, identification, taxonomy, and ecology, and people with these skills are rare and decreasing in numbers (Birks & Birks, 2000). However, the use of plant macrofossil analysis continues to develop. Together with artifacts made of plant material, macrofossils are the basis for palaeoethnobotany and archaeobotany, and they will continue in this important role. In the future, more sophisticated studies of vegetative plant material will extend our knowledge, for example from peat and fen deposits and also from preserved animal (and human) bodies. Knowledge of the representation of vegetation and species by macrofossils is slowly increasing, in both aquatic and terrestrial (e.g. packrat) locations. More modern representation studies are needed from all types of ecosystems, to aid in the interpretation of past macrofossil records.
Plant macrofossils are being increasingly used in multiproxy studies, usually in conjunction with pollen and chironomid analyses. In a multiproxy study, they provide an essential record of vegetation development either in a mire or bog, or in a lake ecosystem (lake plus its catchment). They have shown the rapidity with which aquatic and plants can respond to climate changes during the late-glacial (Birks, 2000) and can demonstrate the dynamics of ecosystem change in this period (Birks & Birks, 2013, 2015, Birks et al., 2012, 2014). One new development is their use in hypothesis testing as at Sägistalsee (Lotter & Birks, 2003). Another new development is the testing of climate-dependent models of vegetation (forest) development through the Holocene driven by independently reconstructed temperature, against the actual forest development reconstructed by macrofossils. Discrepancies help to differentiate between different agencies of vegetation change, namely climate, hydrology, and human impact (Heiri et al., 2006; Schwörer et al., 2016). Plant macrofossils will probably play a pivotal role in these newly developing modeling approaches.
The role of plant macrofossils in radiocarbon chronology will continue to be essential in Quaternary geology and palaeoecology. High-resolution AMS 14C dates around tephra horizons will establish these as time markers wherever the tephras are found, especially in marine cores. Dating of terrestrial and marine macrofossils from the same sediment cores will help to establish the magnitude of the marine reservoir age and how it has varied. This will be enormously useful in correcting the marine 14C chronology especially during the late-glacial climatic transition. The reconstructions of atmospheric CO2 concentrations from stomatal density on fossil leaf cuticles need to be continued, for example to establish detailed CO2 changes during the Late Glacial that are not discerned by ice-core CO2 records, and also to reconstruct CO2 levels in previous Quaternary and older geological periods. There have been various attempts to retrieve plant DNA from fossil material and sediments, and as ancient DNA techniques improve, fossil DNA assemblages may give information about past flora and vegetation or the diet of extinct animals which is not already apparent from the macrofossils themselves (e.g., Brown & Barnes, 2015; Birks & Birks, 2016, Pedersen et al., 2016). Macrofossils can contribute to addressing many of the 50 pressing questions in palaeoecology (Seddon et al., 2014).
Macrofossils from arid and treeless areas, where lake or peat deposits are rare, are perhaps the only means of obtaining a phytogeographic and vegetation history. Plant remains are preserved in rodent middens, alluvial deposits, and also in archaeological contexts in such areas. Macrofossils are also the best means of studying arctic and alpine phytogeography and vegetational development, as the pollen record is fraught with difficulties. Knowledge about these areas of the world, both about the natural vegetation development and also the impact of human activities on it, is steadily increasing through the application of macrofossil analysis. Macrofossil analyses, usually in the context of multidisciplinary studies, are important in evaluating human impact on lake ecosystems, revealing the history of acidification, eutrophication, and pollution and their effects on macrophyte communities and hence on whole aquatic ecosystems. The astonishing rapidity of aquatic-ecosystem responses to human impact, especially in arid lands, has been demonstrated through macrofossil analyses (e.g. Birks et al., 2001a, 2001b), and it is hoped that this signal has been received by organizations concerned with the management of hydrological systems and water sustainability.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780124095489104993
Elicitation
Jonathan Gorelick, Nirit Bernstein, in Advances in Agronomy, 2014
2.2.3.1 UV-B radiation
Elevated UV-B radiation has pleiotropic effects on plant development, morphology, and physiology. The most common protective mechanism against potentially damaging irradiation is the biosynthesis of UV-absorbing compounds (Hahlbrock and Scheel, 1989). These secondary metabolites, mainly phenolic compounds, flavonoids, anthocyanins, and hydroxycinnamate esters, are well known to accumulate in the plant cells and reduce the penetration of the UV-B radiation into deeper cell layers, as well as to detoxify ROS (Caldwell et al., 2007; Kakani et al., 2003). ROSs increase in response to UV-B, as well as salicyclic acid and jasmonic acid, which may all affect the production of secondary metabolites.
It is, therefore, to be expected that in medicinal plants as well, UV radiation stimulates secondary metabolite production. The medicinal plant, Acorus calamus (sweet flag), is used traditionally as a stimulant (Lavrenov and Lavrenova, 1999) and, more recently, it has been shown to have anticarcinogenic activity (Mehrotra et al., 2003). Increasing UV-B exposure in field-grown plants not only increased the total essential oil and phenolics content but also decreased the amount of the possibly toxic beta-asarone (Kumari et al., 2009). These findings are to be expected as phenolics are known UV protectants (Rozema et al., 2002). Increased UV-B exposure also increased the glycyrrhizin content in liquorice roots by almost 50% (Afreen et al., 2005), and increased the accumulation of anthocyanin in the purple top turnip “Tsuda” (Wang et al., 2012).
In addition to its effects on secondary metabolites that are involved in UV shielding and stress protection, UV radiation also elicits increased accumulation of secondary metabolites that are involved in the defense against herbivores and pathogens. It increased benzyl GS content in Tropaeolum majus L. (Schreiner et al., 2009); induced a transient increase in several GSs such as 4-methylsulfinylbutyl GS and GS metabolism-related genes in A. thaliana (Wang et al., 2011); and induced production of GS, especially of 4-methylsulfinylbutyl GS and 4-methoxy-indol-3-ylmethyl GS, in broccoli (Brassica oleracea) (Mewis et al., 2012).
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B978012800138700005X
Drought and nitrogen stress effects and tolerance mechanisms in tomato: a review
J. Machado, … S.M.P. Carvalho, in Plant Nutrition and Food Security in the Era of Climate Change, 2022
12.2.3 Molecular mechanisms underlying drought tolerance
Water deficit imposes selective pressure in the evolution of plant morphology and physiology. Breeding for drought-tolerant crops is a pressing issue due to the increasing frequency and duration of droughts caused by climate change. The activation of molecular pathways modulates the expression level of specific genes responsible for the accumulation of various osmolytes, improvement of the antioxidant system, reduction of transpiration, inhibition of shoot growth, and decrease of tillering, including hormonal regulation (Foolad, 2007; Joshi et al., 2016). Although ABA is reported to be abundant under drought conditions, inducing the expression of various stress-related genes, it has been shown that drought-inducible gene expression is also headed by ABA-independent regulatory systems (Joshi et al., 2016).
The genetic determinants of tomato response to drought are still poorly understood. Many quantitative trait loci (QTL) have been identified (Fooland et al., 2003; Gur and Zamir, 2004). However, the genomic regions covered by the QTL usually cover many genes, limiting the understanding of those involved in drought tolerance. Foolad et al. (2003) identified four drought-tolerant QTLs highly relevant for seed germination under drought stress, while Gur and Zamir (2004) reported three QTLs regulating increased yield under drought conditions. More recently, high-throughput analysis has identified thousands of genes whose expression is altered by drought stress (Candar-Cakir et al., 2016; Iovieno et al., 2016; Liu et al., 2017; Liu et al., 2018), but few studies have deeply analyzed the relevance of downregulated or upregulated genes on tomato drought-stress tolerance improvement (summarized in Table 12.1).
Table 12.1. Summary of genes in which overexpression has been associated with improved tolerance to drought stress in tomato.
| Gene name | Encoded protein | Protein function on drought tolerance | Reference |
| AREB1 | Abscisic acid (ABA)-insensitive-5-like protein |
|
(Orellana et al., 2010) |
| CML44 | Hypothetical calcium-binding protein CML44 |
|
Munir et al. (2016) |
| ERF5 | Ethylene-responsive transcription factor 5 |
|
Pan et al. (2012) |
| HsfA1a | Heat stress transcription factor A-1a |
|
Wang et al. (2015) |
| JUB1 | Transcription factor JUNGBRUNNEN 1 |
|
Thirumalaikumar et al. (2018) |
| MAPK1-3 | Mitogen-activated protein kinases |
|
Li et al. (2013) |
|
Wang et al. (2017) | ||
|
Muhammad et al. (2019) | ||
| MX1 | MIXTA-like MYB transcription factor |
|
Ewas et al. (2016) |
| NPR1 | Regulatory protein NPR1 |
|
Li et al. (2019a) |
| PIP | Aquaporins (plasma membrane intrinsic proteins) |
|
Li et al. (2016) |
| PKE1 |
|
Li et al. (2018); Li et al. (2019b) | |
| SHN1 | Ethylene-responsive transcription factor WIN1 |
|
Al-Abdallat, et al. (2014) |
| WRKY8 | WRKY transcription factor 8 |
|
Gao et al., 2020 |
| ZIP1 | Basic leucine zipper 1 |
|
Zhu et al. (2018) |
After a long and high-pressure selection process mostly based on yield traits, modern tomato cultivars have a very narrow genetic basis, and the vast majority are now drought-stress sensitive (Shirasawa et al., 2010). However, elite genes for stress tolerance appear to be well conserved in some wild tomato species, as Solanum pennellii and accessions from this species show high levels of drought tolerance (Bolger et al., 2014). A model of the different processes and genes involved in the S. pennellii drought tolerance response was proposed by Egea et al. (2018). Several genes linked to different physiological and metabolic processes have been identified, especially those involved in N assimilation, GOGAT/GS cycle, and GABA shunt. An upregulation of genes linked to jasmonic acid/ethylene (JA/ET) biosynthesis was also observed under drought stress in drought-tolerant species. Gong et al. (2010) studied the gene expression of two drought-tolerant introgression lines of S. pennellii, and one drought-sensitive cultivar (S. lycopersicum cultivar ‘M82’) showing that the drought-tolerant lines induced around 400 drought-responsive genes encoding for transcription factors and signaling proteins involved in the cell-wall structure, wax biosynthesis, and plant height. Key signaling molecular pathways regulating gluconeogenesis, purine and pyrimidine nucleotide biosynthesis, tryptophan degradation, starch degradation, methionine biosynthesis, and the removal of superoxide radicals were also specifically affected by drought stress, showing that tomato plants can adapt to drought conditions through decreasing energy dissipation, increasing ATP energy provision, and reducing oxidative damage (Gong et al., 2010). Also, genome-wide studies have identified genetic variation on fruit quality components upon drought stress (Albert et al., 2016). The authors proposed 41 putative candidate genes under three constitutive QTLs and 15 interactive or specific QTLs as being implicated on drought stress response on tomato. Regarding the interactive and specific QTLs, genes related to protein protection (chaperone and heat/cold shock proteins), water and solute transport (aquaporins and other transporters), sugar metabolism (sucrose phosphate synthase and invertases), and hormonal signaling (auxin, gibberellin, and ethylene) were identified (Albert et al., 2016). However, improved genetic tools for construction of knockout mutants on tomato are needed to validate the relevance of such genes on overall drought stress response. Therefore, further studies to stablish genetic determinants of drought stress tolerance in tomato are of utmost importance to globally understand the complex regulatory network(s) controlling this process.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780128229163000147
NONFLORAL HOMOPLASY AND EVOLUTIONARY SCENARIOS IN LIVING AND FOSSIL LAND PLANTS
RICHARD M. Bateman, in Homoplasy, 1996
PROFOUND SHIFTS IN ECOLOGICAL ROLES: AQUATICS AND HETEROTROPHS
Many of the most radical evolutionary simplifications in land plant morphology are associated with profound shifts in ecological roles. The derived, ecologically specialized products of such shifts have attracted preferential attention from morphological and especially molecular phylogeneticists, perhaps in part because their vegetative modification (generally simplification, with the obvious exception of insectivores) has rendered their phylogenetic relationships especially ambiguous.
One such example has already been discussed in the context of the rhizomorphic lycopsids and hydropteridalean water ferns—extreme vegetative reduction and specialization for semiaquatic and aquatic habitats. Common consequences of such changes are reduction or loss of rooting systems (associated with transfer of absorptive functions to modified leaves in the Salviniaceae), decreased proportions of strengthening and/or vascular tissues, elongation and thinning of leaf-bearing axes, webbing of surficial leaves and dissection of subsurficial leaves, modification of stomata and their concentration in parts of the plant still frequently exposed to the atmosphere, and in fully aquatic species the evolution of flotation aids such as internal air spaces and ribs or large trichomes on leaves (e.g., Crawford 1987).
Apart from the aforementioned hydropteridalean ferns (Raubeson et al. 1994; Rothwell and Stockey 1994) and Ceratopteris (Page 1979), the best known examples of aquatic adaptations occur among the angiosperms. In most cases, highly reduced genera have proved to be phylogenetically nested within typically less specialized nonaquatic families. On fossil (G. W. Rothwell personal communication 1995), extant morphological (Mayo 1993) and chloroplast DNA (French et al. 1993) evidence, the tiny floating duckweeds of the genus Lemna (Fig. 10A) are nested within the Araceae; the cryptocorynid aroids (Fig. 10B) form the most likely sister-group to Lemna. Other examples of vegetatively reduced hydrophilic angiosperms include the submerged aquatic Callitriche, which is monophyletic and nested within the Scrophulariaceae (Reeves and Olmstead 1993; Philbrick and Les 1995), and the riparian annual Navarettia, nested within the Polemoniaceae (Spencer 1993).

FIGURE 10. Positive correlations between ecological shifts and parallel radical changes in phenotype. Hydrophily (French et al. 1993; Mayo 1993); (A) the vegetatively reduced aquatic Lemna polyrrhiza, and (B) its nonaquatic aroid sister-group, here illustrated by Arum italicum. Heterotrophy (Williams et al. 1994); (C) the complex, ecologically specialized spring-trap insectivore Dionaea muscipula and (D) its noninsectivorous sister-group in the Plumbaginaceae, here illustrated by Plumbago auriculata.
(A and B after Ross-Craig 1973, Figs. XXXI plate 40B, XXX plate 4; C after Cheers 1992, p. 62; D after Luteyn 1990, Fig. 5)Copyright © 1990
Of greater phylogenetic significance are two groups of vegetatively simple aquatics that have each been implicated on both morphological and molecular evidence as potentially the most primitive extant angiosperms: the Nympheaceae (Hamby and Zimmer 1992) and Ceratophyllum (Les 1988; Crepet et al. 1993; Chase et al. 1993; Endress 1994; Crane et al. 1995). Ceratophyllum, in particular, is still strongly favored as a near-basal angiosperm, implying that its vegetative simplicity could be plesiomorphic rather than secondary within the angiosperms. Resolution of this question is necessary to infer whether the angiosperms could have originated in an aquatic habitat (a similar aquatic–terrestrial transition has been inferred for the apomictic heterosporous pteridophytes and earliest gymnosperms: Bateman and DiMichele 1994b; DiMichele and Bateman 1996). However, Ceratophyllum occupies an extremely long branch of the rbcL tree (e.g., Chase et al. 1993) and its perceived phylogenetic position may therefore be an erroneous case of long–branch attraction (e.g., Hendy and Penny 1989; Donoghue 1994; Penny et al. 1994). Whatever the true nature of the oldest angiosperm (Doyle 1994), transitions between terrestrial and aquatic habitat preferences—together with the associated radical vegetative and life history transitions—have occurred many times during the evolutionary history of the plant kingdom. Similar arguments can be made for iterative origins of tropical mangrove trees and of annual from perennial life histories among angiosperms.
Another suite of radical morphological changes associated with major shifts in ecological roles is the acquisition of at least partially heterotrophic nutrition in previously autotrophic plant lineages.
Saprophytism and parasitism in many phylogenetically disparate ground-dwelling genera (e.g., Orobanche, Monotropa, Rafflesia) generally lead to greatly reduced rooting structures and leaves (typically functionally replaced by various forms of haustoria), and substantial or even complete loss of chlorophyll. Aerial epiphytic parasites such as Viscum are more likely to retain chlorophyllous leaves, but in both cases flowers become the main morphological indicators of phylogenetic relationship. The hemiparasitic and parasitic Scrophulariceae and Orobanchaceae have recently been shown to be collectively monophyletic (DePamphilis and Young 1995; Wolfe and DePamphilis 1995). With this exception, these phenomena have not yet been subjected to systematic phylogenetic surveys; nonetheless, clearly they have evolved independently on many occasions.
An alternative approach to heterotrophic nutrition in plants, especially in nitrogen deficient soils, is insectivory. Unlike all of the case studies discussed above, this life-history transition tends to increase rather than decrease overall morphological complexity (Figs. 10C, 10D). The tremendous range of complex adaptations to insectivory—in particular the various spring, bladder, passive and active flypaper, and pitcher traps, and their underlying physiological specializations—have long fascinated comparative morphologists (e.g., Darwin 1875; Juniper et al. 1989). Recent rbcL phylogenies (Albert et al. 1992; Williams et al. 1994) have identified approximately seven independent evolutionary origins of carnivory; some encompass only one mode of insect capture, but three modes occur in different members of the Droseraceae.
Interestingly, where traditional morphological opinions on the phylogenetic positions of various types of heterotrophs have been tested using molecular data, the original morphological inferences have been largely upheld. Moreover, no clear reversals to autotrophy have been documented in any heterotrophic clade, suggesting that these nutritional adaptations rapidly escalate to inescapable physiological constraints—presumably their loss inevitably causes the death of the deficient individual. Thus, at the coarsest level of resolution of nutritional homologies, present evidence suggests that saprophytism–parasitism and insectivory are both highly iterative and reliably irreversible.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B978012618030550006X
Aphid-Borne Virus Dynamics in the Potato-Weed Pathosystem
Rajagopalbabu Srinivasan, … Juan M. Alvarez, in Insect Pests of Potato, 2013
Effect of Virus-Induced Changes in Potato on Aphid Behavior and Biology
Upon infection, potato viruses induce drastic alterations to potato plant morphology and physiology. These changes are known to affect the behavior and fitness of vectors, and, subsequently, virus spread. Potato viral infections are known to modify host-plant preference of aphids (Castle and Berger 1993, Castle et al. 1998, Srinivasan et al. 2006, Alvarez et al. 2009). The symptoms produced by PLRV-infected potato plants, such as yellowing and stunting, and ensuing physiological changes, arrested winged and wingless aphids longer on PLRV-infected plants than on non-infected potato plants (Castle and Berger 1993, Castle et al. 1998, Eigenbrode et al. 2002, Srinivasan et al. 2006, Alvarez et al. 2009). Studies also have indicated that besides visual and gustatory cues, olfactory cues are associated with aphid preference and/or settling. Volatiles trapped from the headspace of potato plants infected with PLRV more strongly arrested M. persicae nymphs and adults than volatiles trapped from non-infected potato plants (Eigenbrode et al. 2002, Srinivasan et al. 2006, Alvarez et al. 2009).
The settling of aphids on virus-infected plants is more commonly associated with persistent viruses and potato-colonizing aphids. Two potato-colonizing species (M. persicae and M. euphorbiae) consistently settled on PLRV-infected plants more often than on PVY-infected plants. This effect was more prominent when PLRV-infected plants also were infected with PVY (Fig. 11.1). Upon mixed infection, potato plants typically expressed more severe symptoms compared to plants infected with one virus (Srinivasan and Alvarez 2007). It is not uncommon to find plants simultaneously infected with PLRV and PVY in potato ecosystems. The prolonged arrestment of aphids on PLRV-infected plants may provide aphids with a longer acquisition access period, which in turn may aid in successful acquisition and inoculation of the virus.

FIGURE 11.1. Settling of Myzus persicae and Macrosiphum euphorbiae on Potato leafroll virus (PLRV)-infected and Potato virus Y (PVY)-infected potato plants. Alphabets (a–d) indicate differences among treatments. Treatments or bars with the same alphabet are not different from each other.
(With permission: Srinivasan and Alvarez 2007: J. Econ. Entomol. 100: 646–655.)
On the contrary, the effect of non-persistent virus infections on aphid settling behavior has been negligible or minimal. Our studies indicated that winged and wingless morphs of M. persicae and M. euphorbiae did not prefer potato plants infected with PVY to non-infected plants. Unlike persistent viruses, non-persistent viruses are acquired and transmitted by both colonizing and non-colonizing aphids’ in short epidermal probes. Shorter durations of aphid arrestment may favor the spread of non-persistent virus, as their acquisition and inoculation periods typically last only a few seconds. Research conducted on vector behavior as influenced by non-persistent viruses in the potato system and in other systems indicates that virus-infected plants may be arresting aphids for a short period of time to facilitate their spread (Mauck et al. 2010). However, these effects seem to vary between aphid species.
Electrical penetration graph (EPG) studies have shed more light on the effect of non-persistent virus infection on feeding behavior of colonizing and non-colonizing aphids. Boquel et al. (2012) compared feeding behavioral differences of M. euphorbiae on PVY-infected vs. non-infected potato foliage and observed that the duration of the E1 and E2 phases (associated with salivation in the phloem and phloem sap ingestion) decreased and the number of probes increased with PVY infection. M. euphorbiae is a very mobile aphid (Alyokhin and Sewell 2003); therefore, more probing and less time spent in the phloem ingestion phase could likely increase the probability of PVY acquisition and transmission.
In contrast, M. persicae, the most efficient vector of PVY, started probing sooner on PVY-infected plants than on healthy plants. The duration of E2 increased in PVY-infected plants vs non-infected plants. Acquisition and inoculation of a non-persistent virus are associated with subphases II-1 and II-3, respectively, of the potential drop that occurs during a stylet cell puncture (Powell et al. 1995). When comparing the effect of PVY infection on feeding behavior of colonizing vs. non-colonizing aphids, Boquel et al. (2012) observed that the mean duration of the potential drop and that of subphase II-3 increased in PVY-infected plants for M. persicae. On the contrary, for the non-colonizing aphid species Sitobion avenae Fabricius, the mean duration of subphases II-1 and II-3 was reduced on PVY-infected plants.
Besides influencing the behavior of aphids, potato virus infections have also been known to alter the biology of their aphid vectors. As in the case of behavior, the effects induced by a persistent virus seem to positively influence the fitness of colonizing aphids. Vector life history studies conducted with potato or other Solanaceae hosts infected with PLRV revealed that aphids multiplied faster on infected plants than on their non-infected counterparts (Ponsen 1969,Castle and Berger 1993). M. persicae and M. euphorbiae nymphal survival rates in general were greater on PLRV-infected plants than on non-infected plants. The fecundity and intrinsic rate of increase of colonizing aphids were higher on PLRV-infected plants than on non-infected plants (Fig. 11.2) (Srinivasan et al. 2008). Ponsen (1969) also found similar results; the daily fecundity of M. persicae was greater on PLRV-infected P. floridana than on non-infected plants. The reproductive period of M. persicae and M. euphorbiae on PLRV-infected potato plants was longer than on non-infected potato plants (Srinivasan et al. 2008). The exact physiological basis of the responses observed in these studies is unknown, but it is widely speculated that the improved nutritional quality, reduced concentrations of deterrents, and the presence and absence of feeding stimulants and toxicants, respectively, in the phloem of PLRV-infected plants could have positively influenced the fitness of aphids (Guntner et al. 1997, Karley 2002). The interactions between persistently transmitted luteoviruses, such as PLRV, and their vectors are postulated to be mutualistic and may have co-evolved, leading to increased fitness of the vector and propagation of the virus (Castle and Berger 1993, Srinivasan et al. 2008).

FIGURE 11.2. Fecundity, intrinsic rate of increase and longevity of Myzus persicae and Macrosiphum euphorbiae on Potato leafroll virus (PLRV)-infected and non—infected potato plants.
On the contrary, positive effects of non-persistent viruses on vector fitness are not consistent. The fecundity of M. persicae was greater on PVY-infected plants than on non-infected potato plants. However, the fecundity on PVY-infected plants was less than on PLRV-infected plants (Fig. 11.3) (Srinivasan and Alvarez 2007). In the case of M. euphorbiae, no difference in fecundity was observed between PVY-infected and non-infected potato plants. However, significant increase in fecundity was observed on PLRV-infected plants compared to PVY-infected or non-infected potato plants (Fig. 11.3) (Srinivasan and Alvarez 2007).

FIGURE 11.3. Fecundity of Myzus persicae and Macrosiphum euphorbiae on Potato leafroll virus (PLRV)-infected and Potato virus Y (PVY)—infected potato plants. Alphabets (a–d) indicate differences among treatments. Treatments or bars with the same alphabet are not different from each other.
(With permission: Srinivasan and Alvarez 2007: J. Econ. Entomol. 100: 646–655.)
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780123868954000119
Integrated Views in Plant Breeding
Grazia M. Borrelli, … Luigi Cattivelli, in Crop Physiology, 2009
4.2.3 Morphological and developmental modifications
Plant morphology and architecture and the transcriptional networks controlling plant development have been targeted in yield improvement studies. Examples concern the modification of root growth, by affecting cell expansion, and of leaf and stem architecture, by acting on gibberellin’s and brassinosteroid’s signalling pathways (Van Camp, 2005). The identification of the genes responsible for plant dwarfing, related to plant response to gibberellin, or affecting the formation of tillers and lateral branches carrying inflorescences has opened promising ways for yield improvement (Takeda et al., 2003; Sakamoto and Matsuoka, 2004). The alteration of phytochrome phyA and phyB gene expression also represents a powerful strategy to alter plant morphology. By means of phytochrome photoreceptors, plants sense the reduction in the ratio of red (R) to far-red (FR) light when they are shaded by their neighbours and respond by increasing their stem extension, reducing branching, accelerating leaf senescence and reducing allocation of resources to yield organs (Smith, 2000; Libenson et al., 2002). Transgenic overexpression of oat phyA in tobacco showed an enhanced allocation of assimilates to leaves with a consequent increase in fresh leaf harvest index, particularly when plants were grown at high density in the field (Robson et al., 1996). Positive effects on leaf morphology, photosynthetic performance and tuber yield were found in transgenic potatoes overexpressing Arabidopsis phyB (Thiele et al., 1999; Schittenhelm et al., 2004) that were confirmed in field studies at high plant population density (Boccalandro et al., 2003). The enhanced phyB expression not only altered the ability of plants to respond to light signals, but also modified the light environment itself, resulting in higher maximum photosynthesis in leaves of all strata of the canopy, largely due to increased leaf stomatal conductance, and a yield increase due to the increased tuber number.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780123744319000141
Moth bean
Ramavtar Sharma, … Man Mohan Sundria, in The Beans and the Peas, 2021
4.3 Biology and crop phenology
The plant morphology differs considerably in different genotypes of moth bean (Fig. 4.1). With short internodes, the partially angular stem grows upto 30 cm. Dense-packed branched moth bean plant can reach a span of up to 150 cm. Both stem and branches are striated with long, spreading and deflexed hairs. The leaflets of trifoliated leaves are variable in size and shape and more or less deeply lobed. Lobes are oblong, acute or acuminate, more or less hairy when young but become glabrescent with age. The inflorescence is raceme on axillary 5–6 cm long peduncle with yellowish hairs. Racemes are capitated with setaceous and ciliated tips protruding beyond the buds (Brink and Jansen, 2006). Bisexual flowers are papilionaceous and the calyx is campanulated and 4–5 mm long. Being a member of subgenera Ceratotropis (Asiatic Vigna), moth bean flowers always exhibits different shades of yellow color (Baudoin and Marechal, 1988). The yellowish corolla is 5–5.5 mm long. The keel is sickle-shaped. Ten stamens are arranged in nine united and one free fashion. With incurved style, the ovary is sessile and superior. Fruits are cylindrical pods (2.4 × 0.5 cm) having curved beak covered with short stiff hairs. Pod color is brown or pale gray when mature and has 5–8 seed. Rectangular to cylindrical seeds, occasionally reniform shaped, 4.5 × 3 mm are whitish-green, yellow to brown often mottled with black. The linear whitish hilum is nonprotruding. Epigeal germination is rule in moth bean (Majumdar, 2011).
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780128214503000044
THE WORLD OF FOOD GRAINS
J. Wyatt, in Encyclopedia of Food Grains (Second Edition), 2016
Introduction
The definition of plant morphology is the physical appearance of a plant. It can be applied to any species and involves a detailed study of vegetative and reproductive characters in order to form a profile of a plant, which can be used to make general comparisons of plant species displaying a similar structure or detailed comparisons within a species to identify varieties (cultivars). Identifying varieties and species of plants or grains by morphological means is still the main method used in cereal breeding and varietal identification and provides the basis for the definitive description of a variety.
Plants within a species share a similar genetic base, and thus, it is possible to group them into a taxonomic family. ‘Cereals’ is a generic term used to encompass the major grain-producing species whose products are some of the world’s most important staple foods and have been since man started growing them for food some 12 000 years ago. A cereal variety is a population of plants that can be identified as being clearly distinguishable from another population by stable inherited morphological characters. Wheat (Triticum aestivum), rice (Oryza sativa), and maize (Zea mays) account for over half of the cereals consumed worldwide. Cereals sit in the family Poaceae (synonymous with Gramineae). While cereals differ in detail in the way the characters of each species are expressed, they share a general commonality in growth habit and flowering parts. By understanding the morphology of each species, it is possible to use it to identify the varieties (cultivars) that exist in each classification. Recognition of the importance attached to identifying varieties is highlighted by the existence of UPOV (The International Union for the Protection of New Plant Varieties). The test guidelines put together by the 71 members of UPOV provide useful information of the major characters used in establishing the morphological identification of the different varieties of cereal species.
The following section begins with a description of the main morphological characters common to all the cereal species and is followed by further sections specific to each species. Illustrations and photographs are used to help in the identification of the characters described.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780123944375000097
Trichoderma-plant-pathogen interactions for benefit of agriculture and environment
Narendra Kumar, S.M. Paul Khurana, in Biocontrol Agents and Secondary Metabolites, 2021
3.3 Effect on plant physiology, effect on yield and quality of produce
The Trichoderma spp. have the power to changing the plant physiology, viz., net photosynthetic rate, stomatal conductance, internal CO2 concentration, transpiration, nutrient uptake, and water use efficiency. Thus, nutrient uptake enhances physiological processes in plants that lead to good growth (Doni et al., 2014). Micronutrients such as Nitrogen and Magnesium are the components of chlorophyll and enzymes of m-RNA synthesis and gene regulation engaged in photosynthesis. Trichoderma harzianum significantly increased the ability of rice plants to tolerate drought stress and increase in rice plant’s water-holding capacity (Shukla et al., 2012). An approximately threefold increase in the net photosynthetic rate and stomatal conductance and twofold increase in water use efficiency in Trichoderma-treated rice plants as compared to NPK-treated plants has been observed (Doni et al., 2014). The increased photosynthetic rates with less transpiration in Trichoderma treats projects efficiency of high water use (Doni et al., 2014). The high photosynthetic potential in Trichoderma-treated plants with less CO2 concentration can be correlated as an activity of carboxylation through CO2 fixation for glucose production at the time of carbohydrate metabolism, which is very active (Thakur et al., 2010). The prolonged photosynthetic activity supporting delayed senescence in rice is due to activity of Trichoderma spp. (Mishra and Salokhe, 2011). Trichoderma releases cellulases which degrade cellulose and enhance the organic matter and nutrients in the rhizosphere. Jiang et al. (2011) reported that increase of plant physiological processes such as solubilization and minerals chelation may enhance nutrient availability that can upgrade plant metabolism (Harman et al., 2004).
3.3.1 Effect on yield and quality of produce
Trichoderma effect on seed germination, plant morphology, and physiology leads to a better field stand and it also accelerates the vegetative and reproductive growth of plants. It enhances the number of branches, spikes, flowers, and fruits per plant. In many cases, the average weight of individual fruit is also comparatively higher. Higher yield by the application of Trichoderma species in mustard, wheat, corn, tuberose, sugarcane, tomato, okra, etc. has been found (Haque et al., 2012; El-Katatny and Idres, 2014; Naznin et al., 2015; Idowu et al., 2016).
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B978012822919400003X
Inter- and Intraspecific Morphological Variation in Bumblebee Species, and Competition in Flower Utilization
Tamiji Inoue, Makoto Kato, in Effects of Resource Distribution on Animal–Plant Interactions, 1992
C Plant Characters That Relate to Pollination
From the viewpoint of plants, plant morphology is generally thought to affect fertilization success, by increasing pollinator visitation and by decreasing improper pollen transfer (Rathcke, 1983). Visitation intensity of insects is basically determined by the attractiveness of plants (Chapter 5), or the net energetic intake from the plant per unit time (Pleasants, 1980). Rare plants produce much more nectar per flower than do abundant plants to increase their attractiveness (Heinrich, 1976a, b, c; Pleasants, 1980). Flower morphology adds a level of specificity to attractiveness by excluding noneffective pollinators, or robbers. Long flowers protect nectar from short-tongued bees and, as exemplified by Rabdosia longituba, characters other than flower depth may also determine the accessibility of flowers to bees. Such exclusion mechanisms have been thought not to be present in short and dish-shaped flowers, but this is not necessarily true (Kakutani et al., 1989). Short flowers of the Japanese orchid, Platanthera nipponica, for example, that are pollinated by short-tongued noctuids, exclude long-tongued sphingids by having twisted spurs, because only straight spurs can be probed by the long tongues of hovering sphingids (Inoue, 1983; another example in Impatiens in Kato et al., 1991). Neither type can escape from piercing by primary nectar robbers (Inouye, 1980; Roubik, 1989, 1990, see Chapter 11).
Intraspecific size variation is also found in flowers, and different-sized flowers within a single plant species attract different-sized bees. Larger flowers of cow vetch, Vicia cracca, were visited by larger workers of Bombus vagans, and vice versa (Morse, 1978). Assortive mating among similar size classes was found in this plant. Galen and Newport (1987) investigated optimal flower design in sky pilot, Polemonium viscosum. Larger flowers were advantageous because they increased the number of visits by pollinators (higher reward in large flowers), but suffered from a lower frequency of precise pollen transfer. Optimization of flower size results from stabilizing selection in this plant, which balances the opposing forces. By changing spur lengths of orchids experimentally, Inoue (1987) found that shortened spurs decreased both pollinia removal rate (paternal success) and fertilization rate (maternal success). There were no significant deleterious effects of elongating spurs and, in this case, longer flowers would be favored. Our study did not deal with intraspecific size variation of plants. We suggest that simultaneous studies of size variation of both plants and bees would prove fruitful.
The positions at which pollen or pollinia are attached to pollinator bodies are determined by flower morphology. Sharing of a single pollinator by multiple plant species becomes possible if pollen deposition sites vary among the plants (Waser, 1983; Macior, 1982; Feinsinger, 1987; Kato et al., 1991). Sharing pollinators in this way is especially important for rare plants (Rathcke, 1983) to receive the service of trapliners (Janzen, 1971).
However, inter- and intraspecific morphological variation among plants does not always imply specialization of pollinator species specific to individual plant species or morphs (Chapter 11). There was considerable interspecific variation, for example, in the flower morphology of North American Pedicularis, but each species was visited by several common bumblebee species (Macior, 1982, but he did not measure bee morphology). Even specific morphological characters that seem specialized for a single pollinator are frequently pillaged by robbers, parasites, and others (Roubik, 1989; Chapter 11).
Color and odor are other floral characters that advertise flowers to pollinators (Keven, 1978; Macior, 1978). Intraspecific color morphs can promote assortive matings (Brown and Clegg, 1984), while intraspecific odor morphs, each of which is pollinated differentially by bumblebees and flies, are also known (Galen et al., 1987). The classification of pollinator syndromes based on color and odor is found in classical works of pollination ecology.
Flowering phenology has been studied frequently in relation to competition for pollination (Rathcke, 1983). Both staggering of flowering periods among plant members in the same guild that share a single pollinator (Pleasants, 1980; Heinrich, 1976b; Yumoto, 1986, 1987), and random associations (Rathcke, 1984; Tepedino and Stanton, 1981, 1982) are reported from different communities. Such wide variation among the results, which depends on the communities studied and the researchers’ prejudice, demands more careful examination of the patterns. For this purpose, methodologies for the statistical testing of data with null models have been developed by various authors (Pleasants, 1980; Rathcke, 1984). Pleasants (1990) examined by several methods the flowering times of plant species that were hypothetically staggered, and found that his original analysis of the plant–bee communities in the Rocky Mountains (Pleasants, 1980), by mean pairwise overlap, was statistically correct. But in Pleasants (1980), the problem is not in the statistical analysis, but in the rather arbitrary procedure used to assign plants to specific guilds. Boundaries between guilds are generally not distinct, but gradual, even in Pleasants (1980) data. We need a statistically rational standard to discriminate among guilds (Jaksic and Medel, 1990). Such a classification of plants according to pollinator syndromes tends, in many cases, to exaggerate specialization (Waser, 1983).
In the present study, we used cluster analysis for the classification of plants by visitation, but we could not detect distinct guilds of plants visited by four out of five bumblebee species. As shown below, in combinations of two or three plant species, overlap of flowering periods decreases seed set (see Chapter 5 for a further discussion of apparent competition among plants for pollinators generated by differences in nectar production rates). At the community level, however, staggering of flowering periods among plant species appears rare, and is incomplete if present. In addition, the phylogeny of plants (Kochmer and Handel, 1986) and habitat conditions (Macior, 1983) limit the free allocation of flowering periods by plants that is a basic assumption of null models. Simultaneous analyses of both morphology and phenology have been suggested (Murray et al., 1987).
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780080918815500165
Plant Macrofossil Introduction☆
H.H. Birks, in Reference Module in Earth Systems and Environmental Sciences, 2017
How Will Plant Macrofossil Analysis Develop in the Future?
Macrofossil analysis requires good botanical knowledge of plant morphology, identification, taxonomy, and ecology, and people with these skills are rare and decreasing in numbers (Birks & Birks, 2000). However, the use of plant macrofossil analysis continues to develop. Together with artifacts made of plant material, macrofossils are the basis for palaeoethnobotany and archaeobotany, and they will continue in this important role. In the future, more sophisticated studies of vegetative plant material will extend our knowledge, for example from peat and fen deposits and also from preserved animal (and human) bodies. Knowledge of the representation of vegetation and species by macrofossils is slowly increasing, in both aquatic and terrestrial (e.g. packrat) locations. More modern representation studies are needed from all types of ecosystems, to aid in the interpretation of past macrofossil records.
Plant macrofossils are being increasingly used in multiproxy studies, usually in conjunction with pollen and chironomid analyses. In a multiproxy study, they provide an essential record of vegetation development either in a mire or bog, or in a lake ecosystem (lake plus its catchment). They have shown the rapidity with which aquatic and plants can respond to climate changes during the late-glacial (Birks, 2000) and can demonstrate the dynamics of ecosystem change in this period (Birks & Birks, 2013, 2015, Birks et al., 2012, 2014). One new development is their use in hypothesis testing as at Sägistalsee (Lotter & Birks, 2003). Another new development is the testing of climate-dependent models of vegetation (forest) development through the Holocene driven by independently reconstructed temperature, against the actual forest development reconstructed by macrofossils. Discrepancies help to differentiate between different agencies of vegetation change, namely climate, hydrology, and human impact (Heiri et al., 2006; Schwörer et al., 2016). Plant macrofossils will probably play a pivotal role in these newly developing modeling approaches.
The role of plant macrofossils in radiocarbon chronology will continue to be essential in Quaternary geology and palaeoecology. High-resolution AMS 14C dates around tephra horizons will establish these as time markers wherever the tephras are found, especially in marine cores. Dating of terrestrial and marine macrofossils from the same sediment cores will help to establish the magnitude of the marine reservoir age and how it has varied. This will be enormously useful in correcting the marine 14C chronology especially during the late-glacial climatic transition. The reconstructions of atmospheric CO2 concentrations from stomatal density on fossil leaf cuticles need to be continued, for example to establish detailed CO2 changes during the Late Glacial that are not discerned by ice-core CO2 records, and also to reconstruct CO2 levels in previous Quaternary and older geological periods. There have been various attempts to retrieve plant DNA from fossil material and sediments, and as ancient DNA techniques improve, fossil DNA assemblages may give information about past flora and vegetation or the diet of extinct animals which is not already apparent from the macrofossils themselves (e.g., Brown & Barnes, 2015; Birks & Birks, 2016, Pedersen et al., 2016). Macrofossils can contribute to addressing many of the 50 pressing questions in palaeoecology (Seddon et al., 2014).
Macrofossils from arid and treeless areas, where lake or peat deposits are rare, are perhaps the only means of obtaining a phytogeographic and vegetation history. Plant remains are preserved in rodent middens, alluvial deposits, and also in archaeological contexts in such areas. Macrofossils are also the best means of studying arctic and alpine phytogeography and vegetational development, as the pollen record is fraught with difficulties. Knowledge about these areas of the world, both about the natural vegetation development and also the impact of human activities on it, is steadily increasing through the application of macrofossil analysis. Macrofossil analyses, usually in the context of multidisciplinary studies, are important in evaluating human impact on lake ecosystems, revealing the history of acidification, eutrophication, and pollution and their effects on macrophyte communities and hence on whole aquatic ecosystems. The astonishing rapidity of aquatic-ecosystem responses to human impact, especially in arid lands, has been demonstrated through macrofossil analyses (e.g. Birks et al., 2001a, 2001b), and it is hoped that this signal has been received by organizations concerned with the management of hydrological systems and water sustainability.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780124095489104993
Elicitation
Jonathan Gorelick, Nirit Bernstein, in Advances in Agronomy, 2014
2.2.3.1 UV-B radiation
Elevated UV-B radiation has pleiotropic effects on plant development, morphology, and physiology. The most common protective mechanism against potentially damaging irradiation is the biosynthesis of UV-absorbing compounds (Hahlbrock and Scheel, 1989). These secondary metabolites, mainly phenolic compounds, flavonoids, anthocyanins, and hydroxycinnamate esters, are well known to accumulate in the plant cells and reduce the penetration of the UV-B radiation into deeper cell layers, as well as to detoxify ROS (Caldwell et al., 2007; Kakani et al., 2003). ROSs increase in response to UV-B, as well as salicyclic acid and jasmonic acid, which may all affect the production of secondary metabolites.
It is, therefore, to be expected that in medicinal plants as well, UV radiation stimulates secondary metabolite production. The medicinal plant, Acorus calamus (sweet flag), is used traditionally as a stimulant (Lavrenov and Lavrenova, 1999) and, more recently, it has been shown to have anticarcinogenic activity (Mehrotra et al., 2003). Increasing UV-B exposure in field-grown plants not only increased the total essential oil and phenolics content but also decreased the amount of the possibly toxic beta-asarone (Kumari et al., 2009). These findings are to be expected as phenolics are known UV protectants (Rozema et al., 2002). Increased UV-B exposure also increased the glycyrrhizin content in liquorice roots by almost 50% (Afreen et al., 2005), and increased the accumulation of anthocyanin in the purple top turnip “Tsuda” (Wang et al., 2012).
In addition to its effects on secondary metabolites that are involved in UV shielding and stress protection, UV radiation also elicits increased accumulation of secondary metabolites that are involved in the defense against herbivores and pathogens. It increased benzyl GS content in Tropaeolum majus L. (Schreiner et al., 2009); induced a transient increase in several GSs such as 4-methylsulfinylbutyl GS and GS metabolism-related genes in A. thaliana (Wang et al., 2011); and induced production of GS, especially of 4-methylsulfinylbutyl GS and 4-methoxy-indol-3-ylmethyl GS, in broccoli (Brassica oleracea) (Mewis et al., 2012).
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B978012800138700005X
Drought and nitrogen stress effects and tolerance mechanisms in tomato: a review
J. Machado, … S.M.P. Carvalho, in Plant Nutrition and Food Security in the Era of Climate Change, 2022
12.2.3 Molecular mechanisms underlying drought tolerance
Water deficit imposes selective pressure in the evolution of plant morphology and physiology. Breeding for drought-tolerant crops is a pressing issue due to the increasing frequency and duration of droughts caused by climate change. The activation of molecular pathways modulates the expression level of specific genes responsible for the accumulation of various osmolytes, improvement of the antioxidant system, reduction of transpiration, inhibition of shoot growth, and decrease of tillering, including hormonal regulation (Foolad, 2007; Joshi et al., 2016). Although ABA is reported to be abundant under drought conditions, inducing the expression of various stress-related genes, it has been shown that drought-inducible gene expression is also headed by ABA-independent regulatory systems (Joshi et al., 2016).
The genetic determinants of tomato response to drought are still poorly understood. Many quantitative trait loci (QTL) have been identified (Fooland et al., 2003; Gur and Zamir, 2004). However, the genomic regions covered by the QTL usually cover many genes, limiting the understanding of those involved in drought tolerance. Foolad et al. (2003) identified four drought-tolerant QTLs highly relevant for seed germination under drought stress, while Gur and Zamir (2004) reported three QTLs regulating increased yield under drought conditions. More recently, high-throughput analysis has identified thousands of genes whose expression is altered by drought stress (Candar-Cakir et al., 2016; Iovieno et al., 2016; Liu et al., 2017; Liu et al., 2018), but few studies have deeply analyzed the relevance of downregulated or upregulated genes on tomato drought-stress tolerance improvement (summarized in Table 12.1).
Table 12.1. Summary of genes in which overexpression has been associated with improved tolerance to drought stress in tomato.
| Gene name | Encoded protein | Protein function on drought tolerance | Reference |
| AREB1 | Abscisic acid (ABA)-insensitive-5-like protein |
|
(Orellana et al., 2010) |
| CML44 | Hypothetical calcium-binding protein CML44 |
|
Munir et al. (2016) |
| ERF5 | Ethylene-responsive transcription factor 5 |
|
Pan et al. (2012) |
| HsfA1a | Heat stress transcription factor A-1a |
|
Wang et al. (2015) |
| JUB1 | Transcription factor JUNGBRUNNEN 1 |
|
Thirumalaikumar et al. (2018) |
| MAPK1-3 | Mitogen-activated protein kinases |
|
Li et al. (2013) |
|
Wang et al. (2017) | ||
|
Muhammad et al. (2019) | ||
| MX1 | MIXTA-like MYB transcription factor |
|
Ewas et al. (2016) |
| NPR1 | Regulatory protein NPR1 |
|
Li et al. (2019a) |
| PIP | Aquaporins (plasma membrane intrinsic proteins) |
|
Li et al. (2016) |
| PKE1 |
|
Li et al. (2018); Li et al. (2019b) | |
| SHN1 | Ethylene-responsive transcription factor WIN1 |
|
Al-Abdallat, et al. (2014) |
| WRKY8 | WRKY transcription factor 8 |
|
Gao et al., 2020 |
| ZIP1 | Basic leucine zipper 1 |
|
Zhu et al. (2018) |
After a long and high-pressure selection process mostly based on yield traits, modern tomato cultivars have a very narrow genetic basis, and the vast majority are now drought-stress sensitive (Shirasawa et al., 2010). However, elite genes for stress tolerance appear to be well conserved in some wild tomato species, as Solanum pennellii and accessions from this species show high levels of drought tolerance (Bolger et al., 2014). A model of the different processes and genes involved in the S. pennellii drought tolerance response was proposed by Egea et al. (2018). Several genes linked to different physiological and metabolic processes have been identified, especially those involved in N assimilation, GOGAT/GS cycle, and GABA shunt. An upregulation of genes linked to jasmonic acid/ethylene (JA/ET) biosynthesis was also observed under drought stress in drought-tolerant species. Gong et al. (2010) studied the gene expression of two drought-tolerant introgression lines of S. pennellii, and one drought-sensitive cultivar (S. lycopersicum cultivar ‘M82’) showing that the drought-tolerant lines induced around 400 drought-responsive genes encoding for transcription factors and signaling proteins involved in the cell-wall structure, wax biosynthesis, and plant height. Key signaling molecular pathways regulating gluconeogenesis, purine and pyrimidine nucleotide biosynthesis, tryptophan degradation, starch degradation, methionine biosynthesis, and the removal of superoxide radicals were also specifically affected by drought stress, showing that tomato plants can adapt to drought conditions through decreasing energy dissipation, increasing ATP energy provision, and reducing oxidative damage (Gong et al., 2010). Also, genome-wide studies have identified genetic variation on fruit quality components upon drought stress (Albert et al., 2016). The authors proposed 41 putative candidate genes under three constitutive QTLs and 15 interactive or specific QTLs as being implicated on drought stress response on tomato. Regarding the interactive and specific QTLs, genes related to protein protection (chaperone and heat/cold shock proteins), water and solute transport (aquaporins and other transporters), sugar metabolism (sucrose phosphate synthase and invertases), and hormonal signaling (auxin, gibberellin, and ethylene) were identified (Albert et al., 2016). However, improved genetic tools for construction of knockout mutants on tomato are needed to validate the relevance of such genes on overall drought stress response. Therefore, further studies to stablish genetic determinants of drought stress tolerance in tomato are of utmost importance to globally understand the complex regulatory network(s) controlling this process.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780128229163000147
NONFLORAL HOMOPLASY AND EVOLUTIONARY SCENARIOS IN LIVING AND FOSSIL LAND PLANTS
RICHARD M. Bateman, in Homoplasy, 1996
PROFOUND SHIFTS IN ECOLOGICAL ROLES: AQUATICS AND HETEROTROPHS
Many of the most radical evolutionary simplifications in land plant morphology are associated with profound shifts in ecological roles. The derived, ecologically specialized products of such shifts have attracted preferential attention from morphological and especially molecular phylogeneticists, perhaps in part because their vegetative modification (generally simplification, with the obvious exception of insectivores) has rendered their phylogenetic relationships especially ambiguous.
One such example has already been discussed in the context of the rhizomorphic lycopsids and hydropteridalean water ferns—extreme vegetative reduction and specialization for semiaquatic and aquatic habitats. Common consequences of such changes are reduction or loss of rooting systems (associated with transfer of absorptive functions to modified leaves in the Salviniaceae), decreased proportions of strengthening and/or vascular tissues, elongation and thinning of leaf-bearing axes, webbing of surficial leaves and dissection of subsurficial leaves, modification of stomata and their concentration in parts of the plant still frequently exposed to the atmosphere, and in fully aquatic species the evolution of flotation aids such as internal air spaces and ribs or large trichomes on leaves (e.g., Crawford 1987).
Apart from the aforementioned hydropteridalean ferns (Raubeson et al. 1994; Rothwell and Stockey 1994) and Ceratopteris (Page 1979), the best known examples of aquatic adaptations occur among the angiosperms. In most cases, highly reduced genera have proved to be phylogenetically nested within typically less specialized nonaquatic families. On fossil (G. W. Rothwell personal communication 1995), extant morphological (Mayo 1993) and chloroplast DNA (French et al. 1993) evidence, the tiny floating duckweeds of the genus Lemna (Fig. 10A) are nested within the Araceae; the cryptocorynid aroids (Fig. 10B) form the most likely sister-group to Lemna. Other examples of vegetatively reduced hydrophilic angiosperms include the submerged aquatic Callitriche, which is monophyletic and nested within the Scrophulariaceae (Reeves and Olmstead 1993; Philbrick and Les 1995), and the riparian annual Navarettia, nested within the Polemoniaceae (Spencer 1993).
FIGURE 10. Positive correlations between ecological shifts and parallel radical changes in phenotype. Hydrophily (French et al. 1993; Mayo 1993); (A) the vegetatively reduced aquatic Lemna polyrrhiza, and (B) its nonaquatic aroid sister-group, here illustrated by Arum italicum. Heterotrophy (Williams et al. 1994); (C) the complex, ecologically specialized spring-trap insectivore Dionaea muscipula and (D) its noninsectivorous sister-group in the Plumbaginaceae, here illustrated by Plumbago auriculata.
(A and B after Ross-Craig 1973, Figs. XXXI plate 40B, XXX plate 4; C after Cheers 1992, p. 62; D after Luteyn 1990, Fig. 5)Copyright © 1990
Of greater phylogenetic significance are two groups of vegetatively simple aquatics that have each been implicated on both morphological and molecular evidence as potentially the most primitive extant angiosperms: the Nympheaceae (Hamby and Zimmer 1992) and Ceratophyllum (Les 1988; Crepet et al. 1993; Chase et al. 1993; Endress 1994; Crane et al. 1995). Ceratophyllum, in particular, is still strongly favored as a near-basal angiosperm, implying that its vegetative simplicity could be plesiomorphic rather than secondary within the angiosperms. Resolution of this question is necessary to infer whether the angiosperms could have originated in an aquatic habitat (a similar aquatic–terrestrial transition has been inferred for the apomictic heterosporous pteridophytes and earliest gymnosperms: Bateman and DiMichele 1994b; DiMichele and Bateman 1996). However, Ceratophyllum occupies an extremely long branch of the rbcL tree (e.g., Chase et al. 1993) and its perceived phylogenetic position may therefore be an erroneous case of long–branch attraction (e.g., Hendy and Penny 1989; Donoghue 1994; Penny et al. 1994). Whatever the true nature of the oldest angiosperm (Doyle 1994), transitions between terrestrial and aquatic habitat preferences—together with the associated radical vegetative and life history transitions—have occurred many times during the evolutionary history of the plant kingdom. Similar arguments can be made for iterative origins of tropical mangrove trees and of annual from perennial life histories among angiosperms.
Another suite of radical morphological changes associated with major shifts in ecological roles is the acquisition of at least partially heterotrophic nutrition in previously autotrophic plant lineages.
Saprophytism and parasitism in many phylogenetically disparate ground-dwelling genera (e.g., Orobanche, Monotropa, Rafflesia) generally lead to greatly reduced rooting structures and leaves (typically functionally replaced by various forms of haustoria), and substantial or even complete loss of chlorophyll. Aerial epiphytic parasites such as Viscum are more likely to retain chlorophyllous leaves, but in both cases flowers become the main morphological indicators of phylogenetic relationship. The hemiparasitic and parasitic Scrophulariceae and Orobanchaceae have recently been shown to be collectively monophyletic (DePamphilis and Young 1995; Wolfe and DePamphilis 1995). With this exception, these phenomena have not yet been subjected to systematic phylogenetic surveys; nonetheless, clearly they have evolved independently on many occasions.
An alternative approach to heterotrophic nutrition in plants, especially in nitrogen deficient soils, is insectivory. Unlike all of the case studies discussed above, this life-history transition tends to increase rather than decrease overall morphological complexity (Figs. 10C, 10D). The tremendous range of complex adaptations to insectivory—in particular the various spring, bladder, passive and active flypaper, and pitcher traps, and their underlying physiological specializations—have long fascinated comparative morphologists (e.g., Darwin 1875; Juniper et al. 1989). Recent rbcL phylogenies (Albert et al. 1992; Williams et al. 1994) have identified approximately seven independent evolutionary origins of carnivory; some encompass only one mode of insect capture, but three modes occur in different members of the Droseraceae.
Interestingly, where traditional morphological opinions on the phylogenetic positions of various types of heterotrophs have been tested using molecular data, the original morphological inferences have been largely upheld. Moreover, no clear reversals to autotrophy have been documented in any heterotrophic clade, suggesting that these nutritional adaptations rapidly escalate to inescapable physiological constraints—presumably their loss inevitably causes the death of the deficient individual. Thus, at the coarsest level of resolution of nutritional homologies, present evidence suggests that saprophytism–parasitism and insectivory are both highly iterative and reliably irreversible.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B978012618030550006X
Aphid-Borne Virus Dynamics in the Potato-Weed Pathosystem
Rajagopalbabu Srinivasan, … Juan M. Alvarez, in Insect Pests of Potato, 2013
Effect of Virus-Induced Changes in Potato on Aphid Behavior and Biology
Upon infection, potato viruses induce drastic alterations to potato plant morphology and physiology. These changes are known to affect the behavior and fitness of vectors, and, subsequently, virus spread. Potato viral infections are known to modify host-plant preference of aphids (Castle and Berger 1993, Castle et al. 1998, Srinivasan et al. 2006, Alvarez et al. 2009). The symptoms produced by PLRV-infected potato plants, such as yellowing and stunting, and ensuing physiological changes, arrested winged and wingless aphids longer on PLRV-infected plants than on non-infected potato plants (Castle and Berger 1993, Castle et al. 1998, Eigenbrode et al. 2002, Srinivasan et al. 2006, Alvarez et al. 2009). Studies also have indicated that besides visual and gustatory cues, olfactory cues are associated with aphid preference and/or settling. Volatiles trapped from the headspace of potato plants infected with PLRV more strongly arrested M. persicae nymphs and adults than volatiles trapped from non-infected potato plants (Eigenbrode et al. 2002, Srinivasan et al. 2006, Alvarez et al. 2009).
The settling of aphids on virus-infected plants is more commonly associated with persistent viruses and potato-colonizing aphids. Two potato-colonizing species (M. persicae and M. euphorbiae) consistently settled on PLRV-infected plants more often than on PVY-infected plants. This effect was more prominent when PLRV-infected plants also were infected with PVY (Fig. 11.1). Upon mixed infection, potato plants typically expressed more severe symptoms compared to plants infected with one virus (Srinivasan and Alvarez 2007). It is not uncommon to find plants simultaneously infected with PLRV and PVY in potato ecosystems. The prolonged arrestment of aphids on PLRV-infected plants may provide aphids with a longer acquisition access period, which in turn may aid in successful acquisition and inoculation of the virus.
FIGURE 11.1. Settling of Myzus persicae and Macrosiphum euphorbiae on Potato leafroll virus (PLRV)-infected and Potato virus Y (PVY)-infected potato plants. Alphabets (a–d) indicate differences among treatments. Treatments or bars with the same alphabet are not different from each other.
(With permission: Srinivasan and Alvarez 2007: J. Econ. Entomol. 100: 646–655.)
On the contrary, the effect of non-persistent virus infections on aphid settling behavior has been negligible or minimal. Our studies indicated that winged and wingless morphs of M. persicae and M. euphorbiae did not prefer potato plants infected with PVY to non-infected plants. Unlike persistent viruses, non-persistent viruses are acquired and transmitted by both colonizing and non-colonizing aphids’ in short epidermal probes. Shorter durations of aphid arrestment may favor the spread of non-persistent virus, as their acquisition and inoculation periods typically last only a few seconds. Research conducted on vector behavior as influenced by non-persistent viruses in the potato system and in other systems indicates that virus-infected plants may be arresting aphids for a short period of time to facilitate their spread (Mauck et al. 2010). However, these effects seem to vary between aphid species.
Electrical penetration graph (EPG) studies have shed more light on the effect of non-persistent virus infection on feeding behavior of colonizing and non-colonizing aphids. Boquel et al. (2012) compared feeding behavioral differences of M. euphorbiae on PVY-infected vs. non-infected potato foliage and observed that the duration of the E1 and E2 phases (associated with salivation in the phloem and phloem sap ingestion) decreased and the number of probes increased with PVY infection. M. euphorbiae is a very mobile aphid (Alyokhin and Sewell 2003); therefore, more probing and less time spent in the phloem ingestion phase could likely increase the probability of PVY acquisition and transmission.
In contrast, M. persicae, the most efficient vector of PVY, started probing sooner on PVY-infected plants than on healthy plants. The duration of E2 increased in PVY-infected plants vs non-infected plants. Acquisition and inoculation of a non-persistent virus are associated with subphases II-1 and II-3, respectively, of the potential drop that occurs during a stylet cell puncture (Powell et al. 1995). When comparing the effect of PVY infection on feeding behavior of colonizing vs. non-colonizing aphids, Boquel et al. (2012) observed that the mean duration of the potential drop and that of subphase II-3 increased in PVY-infected plants for M. persicae. On the contrary, for the non-colonizing aphid species Sitobion avenae Fabricius, the mean duration of subphases II-1 and II-3 was reduced on PVY-infected plants.
Besides influencing the behavior of aphids, potato virus infections have also been known to alter the biology of their aphid vectors. As in the case of behavior, the effects induced by a persistent virus seem to positively influence the fitness of colonizing aphids. Vector life history studies conducted with potato or other Solanaceae hosts infected with PLRV revealed that aphids multiplied faster on infected plants than on their non-infected counterparts (Ponsen 1969,Castle and Berger 1993). M. persicae and M. euphorbiae nymphal survival rates in general were greater on PLRV-infected plants than on non-infected plants. The fecundity and intrinsic rate of increase of colonizing aphids were higher on PLRV-infected plants than on non-infected plants (Fig. 11.2) (Srinivasan et al. 2008). Ponsen (1969) also found similar results; the daily fecundity of M. persicae was greater on PLRV-infected P. floridana than on non-infected plants. The reproductive period of M. persicae and M. euphorbiae on PLRV-infected potato plants was longer than on non-infected potato plants (Srinivasan et al. 2008). The exact physiological basis of the responses observed in these studies is unknown, but it is widely speculated that the improved nutritional quality, reduced concentrations of deterrents, and the presence and absence of feeding stimulants and toxicants, respectively, in the phloem of PLRV-infected plants could have positively influenced the fitness of aphids (Guntner et al. 1997, Karley 2002). The interactions between persistently transmitted luteoviruses, such as PLRV, and their vectors are postulated to be mutualistic and may have co-evolved, leading to increased fitness of the vector and propagation of the virus (Castle and Berger 1993, Srinivasan et al. 2008).
FIGURE 11.2. Fecundity, intrinsic rate of increase and longevity of Myzus persicae and Macrosiphum euphorbiae on Potato leafroll virus (PLRV)-infected and non—infected potato plants.
On the contrary, positive effects of non-persistent viruses on vector fitness are not consistent. The fecundity of M. persicae was greater on PVY-infected plants than on non-infected potato plants. However, the fecundity on PVY-infected plants was less than on PLRV-infected plants (Fig. 11.3) (Srinivasan and Alvarez 2007). In the case of M. euphorbiae, no difference in fecundity was observed between PVY-infected and non-infected potato plants. However, significant increase in fecundity was observed on PLRV-infected plants compared to PVY-infected or non-infected potato plants (Fig. 11.3) (Srinivasan and Alvarez 2007).
FIGURE 11.3. Fecundity of Myzus persicae and Macrosiphum euphorbiae on Potato leafroll virus (PLRV)-infected and Potato virus Y (PVY)—infected potato plants. Alphabets (a–d) indicate differences among treatments. Treatments or bars with the same alphabet are not different from each other.
(With permission: Srinivasan and Alvarez 2007: J. Econ. Entomol. 100: 646–655.)
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780123868954000119
Курс лекций по морфологии растений дисциплины
ОП.02 «Ботаника» специальности 35.02.01 Лесное и
лесопарковое хозяйство
Лекция
2. Морфология стебля
Новые
понятия и термины: стебель, лист, почка, почечные чешуи,
верхушечная почка; боковые (пазушные), придаточные и спящие почки; почки
возобновления; вегетативная, генеративная и вегетативно — генеративная почки;
почечное кольцо, корневище, клубень, клубнелуковица, луковица, донце, плети
(усы), колючки, усики, суккулентные побеги.
План
1.Определение
побега, стебля
2.Функции
побега и стебля
3.Происхождение
4.Почка.
Ее строение и значение
5.Классификация
почек
6.Видоизменения
побегов и стеблей
7.Характер
расположения стебля в пространстве
Стебель — это осевая часть побега, несущая на себе листья, почки.
Побег
— это стебель с листьями и почками развивающийся в течение одного года.Стебель
— это ось побега, он связывает корни с листьями. Почка — это зачаточный побег.
Функции побега и стебля:
1.Проводящая — связывает подземные и надземные органы (ксилема,
флоэма), осуществляется проводящими тканями стебля и листа
2.Запасающая — накапливает питающие вещества и воду (паренхима)
осуществляется паренхимой стебля и, в некоторых случаях, листа).
3.Вегетативное размножение — за счет образования придаточных почек
и корней (клетки живых тканей);
4.Фотосинтезирующая — как правило, молодые однолетние стебли
(фотосинтезирующая паренхима);
5.Ориентация листьев в пространстве — механические ткани.
Признаки стебля
Радиальная симметрия.
Длительный верхушечный рост.
Длительный рост в ширину.
Характерно повторение однородных участков – метамеров (узел+
междоузлие).
На нем образуются листья и почки.
В центре располагается сердцевина или воздушная полость.
Механические ткани расположены по периферии.
Происхождение
Побег
сформировался в ходе эволюции растений при переходе их к наземному образу
жизни. Сначала он состоял из одних стеблей (как у вегетативных побегов хвощей).
Затем на нем появились листья. Побег развивается из почки.
Почка – зачаточный побег, состоящий из укороченного стебля с
зачаточными листьями. Она покрыта почечными чешуями, плотно прилегающими друг к
другу.
Почечные чешуи – это видоизмененные листья, они защищают почку от
механических повреждений, от холода, излишнего тепла, испарения, от проникновения
болезнетворных бактерий и грибов.
По месту расположения на растении различают следующие типы почек:
1.Верхушечные. Они располагаются на верхушке стебля. Из
верхушечной почки образуется главный побег.
2.Боковые почки — пазушные. Они расположены по бокам стебля, над
местом прикрепления листа, т. е. в пазухе листа. Из боковых почек формируются
боковые побеги, происходит ветвление ,т. е. образование системы побегов.
3.Придаточные почки. Они располагаются на любой части стебля вне
пазухи листа, а также на корнях и листьях. Благодаря придаточным почкам
происходит образование побегов при вегетативном размножении.
Типы
почек по степени их активности:
1.
Почки возобновления
распускаются через год после формирования. Из них ежегодно образуются новые
побеги. Образуются у деревьев, кустарников и многолетних трав.
2.
Спящие почки — особая
категория почек, характерная для деревьев, кустарников и многолетних трав. По
происхождению они пазушные или придаточные. Спящие почки могут «спать» не
развиваться в течение многих лет. Сигналом для «пробуждения» служит повреждение
ствола или ветви.
Типы
почек по функциям:
1.Вегетативная
почка состоит из конуса нарастания стебля, зачатков листьев и зачатков почек.
Она покрыта почечными чешуями.
2.Генеративная почка — цветочная заключает в себе зачаток соцветия
или одиночный цветок, прикрытый почечными чешуями.
3.Вегетативно-генеративная почка заключает в себе зачаточные
листья и почки, а конус нарастания превращен в зачаточный цветок или соцветие.
Образование побега из почки начинается с разрастания листовых зачатков и роста
зачаточного стебля. Наружные почечные чешуи быстро подсыхают и опадают при
начале развертывания почки. От них у основания побега остаются сближенные
рубцы, так называемое почечное кольцо, которое хорошо заметно на ветвях
деревьев и кустарников. По числу почечных колец можно подсчитать возраст ветви.
Побеги, вырастающие из почек за один вегетативный период (весна — лето)
называют годичным приростом.
Видоизменения
побегов
Видоизменения побегов произошли в процессе длительной эволюции, как
приспособления к выполнению специальных функций : запаса питательных веществ,
прикрепления, защиты.
Различают следующие видоизменения побегов:
1.Корневище. Многолетний
подземный побег, имеющий листья в виде бесцветных или бурых чешуи, в пазухах
которых расположены почки. Функции: запас веществ и вегетативное размножение
(ландыш, фиалка, пырей, земляника, брусника, черника).
2.Клубень —
видоизмененный побег с ярко выраженной запасающей функцией стебля. На стебле
хорошо различимы почки. Они располагаются в пазухах чешуевидных листьев. Чешуи
быстро отмирают, и на их месте образуются рубцы. У картофеля почки называются
глазки, а рубцы от чешуевидных листьев — бровки. Клубни могут располагаться над
землей (кольраби) или в почве (картофель, топинамбур). Подземные клубни обычно
образуются на столонах — недолговечных подземных побегах с чешуевидными
листьями. Верхушка этих побегов разрастается, образуя клубень. Функции клубня:
запас питательных веществ и вегетативное размножение (картофель, кольраби,
топинамбур).
3.Клубнелуковица — это облиственный клубень, т. е. клубень, покрытый
чешуевидными сухими листьями и по внешнему виду напоминающий луковицу. Функции:
запас питательных веществ (гладиолус, крокус).
4.Луковица — это укороченный побег, стеблевая часть которого называется донцем.
В луковице различают два типа видоизмененных чешуевидных листьев: листья
сочные, запасающие воду с растворенными питательными веществами, и листья
сухие, покрывающие луковицу снаружи. Из верхушечных и пазушных почек вырастают
зеленые надземные побеги, а на донце образуются придаточные корни. Функции:
запас пита-тельных веществ, вегетативное размножение (лук, лилии, тюльпаны,
нарцисс, чеснок).
5.Надземные столоны (плети) или «усы». Недолговечные ползучие побеги, обычно
лишенные развитых зеленых листьев. Верхушечная почка дает розетку листьев, на
стебле образуются придаточные корни. Функции: вегетативное размножение
(земляника).
6.Колючки — укороченные побеги, имеющие ограниченный рост. На них не образуются
листья и почки. Колючки располагаются на стебле в пазухах листа. Функции:
защита (боярышник, гледичия).
7.Усики — побеги, у которых не образуются листья, а стеблевая часть изгибается,
обвиваясь вокруг опоры. Функция: прикрепление к опоре (виноград, огурец).
8.Суккулентные побеги. Надземные побеги, приспособленные для накопления воды.
Сочный стебель выполняет функцию запаса воды и фотосинтеза, а листья исчезают
или превращаются в колючки (кактусы, молочаи).
Типы
стеблей по характеру расположения в пространстве
1.Прямостоячий стебель расположен вертикально. Способен
самостоятельно удерживать вертикальное положение.
2.Приподнимающийся стебель — сначала растет горизонтально, а затем
в вертикальном положении.
3.Стелющийся стебель растет в горизонтальном направлении,
параллельно поверхности земли. Если стелющийся стебель укореняется, то он
называется ползучим.
4.Вьющийся стебель растет вверх, но ему нужна дополнительная
опора, т. к. в нем плохо развиты механические ткани. Вокруг опоры он и
обвивается.
5.Цепляющийся стебель растет, как и вьющийся, но цепляется за
опору с помощью усиков.
Вопросы на
закрепление:
1.Что называют побегом? Из каких частей он состоит?
2.Какие функции выполняет побег?
3.Каково строение почки? Какие почки Вам известны?
4.С чем связано образование видоизмененных побегов?
5.Чем строение видоизмененных побегов отличается от типичного
побега? 6.В чем отличия друг от друга различных видоизмененных побегов?
7.Какой орган называют стеблем? Какие функции он выполняет?
8.Назовите отличительные черты стебля.
9.Какие типы расположения стебля в пространстве Вам известны?
Лекция
3. Морфология корня
Новые
понятия и термины: корневая система (стержневая,
мочковатая), корни: главный, боковые, придаточные; корневой чехлик, зона
деления, роста всасывания, проведения (ветвления), первичная кора, центральный
цилиндр, пикировка, минеральные и органические удобрения, корнеплод, корнеклубень,
бактериальные клубеньки.
План
1.Определение
корня.
2.Отличительные
признаки корня.
3.Функции
корня.
4.Зоны
корня.
5.Видоизменения
корней
Корень — это осевой вегетативный орган растения. Корень обладает
радиальной симметрией и растет неопределенно долго.
Тело первых вышедших на сушу растений ещё не было расчленено на побеги и корни. Оно
состояло из ответвлений, одни из которых поднимались вертикально, а другие
прижимались к почве и поглощали воду и питательные вещества. Несмотря на
примитивное строение, эти растения были обеспечены водой и питательными
веществами, так как имели небольшие размеры и жили около воды.
В ходе дальнейшей эволюции некоторые ответвления стали углубляться в почву и
дали начало корням, приспособленным к более совершенному почвенному питанию.
Это сопровождалось глубокой перестройкой их структуры и появлением
специализированных тканей. Образование
корней было крупным эволюционным достижением, благодаря которому растения смогли
осваивать более сухие почвы и образовывать крупные побеги, поднятые вверх к
свету. Например, у мохообразных настоящих
корней нет, их вегетативное тело небольших размеров — до 30 см,
обитают мхи во влажных местах. У папоротниковидных появляются
настоящие корни, это приводит к увеличению размеров вегетативного тела и к
расцвету этой группы в каменноугольный период.
Ко́рень (лат. radix) —
осевой, обычно подземный вегетативный орган высших сосудистых растений,
обладающий неограниченным ростом в длину и положительным геотропизмом. Корень
осуществляет закрепление растения в почве и
обеспечивает поглощение и проведение воды с растворёнными минеральными
веществами к стеблю и листьям
На
корне нет листьев, в клетках корня нет хлоропластов.
Кроме основного корня, многие растения имеют боковые и придаточные корни.
Главный корень образуется из зародышевого корешка семени. Боковые
корни отходят от главного, боковых и придаточных. Придаточные корни образуются
на стебле и листьях.
Совокупность
всех корней растения называют корневой системой.
В случае, когда главный корень незначительно выражен, а придаточные корни
выражены значительно, корневая система называется мочковатой (однодольные).
Если главный корень выражен значительно, корневая система называется стержневой (двудольные).
В стержневой корневой системе главный корень сильно развит и
хорошо заметен среди других корней (характерно для двудольных). Разновидность
стержневой корневой системы — ветвистая корневая система:
состоит из нескольких боковых корней, среди которых не различают главный
корень; характерна для деревьев.
В мочковатой
корневой системе на ранних этапах развития главный корень,
образованный зародышевым корешком, отмирает, а корневая система составляется
придаточными корнями (характерна для однодольных). Стержневая корневая система
проникает в почву обычно глубже, чем мочковатая, однако мочковатая корневая
система лучше оплетает прилегающие частицы грунта.
Отличительные
признаки корня
1.Конус
нарастания прикрыт корневым чехликом.
2.Есть
корневые волоски.
3.На
корне никогда не образуются листья, даже чешуевидные.
4.Нет
повторяющихся участков.
Функции
корня
1.Закрепление
растения в почве.
2.Всасывание
воды и минеральных веществ.
3.Проведение
веществ к надземным частям растения.
4.Синтез
веществ (витаминов, гормонов).
5.Запас
веществ.
6.Вегетативное
размножение.
7.
Взаимодействие с корнями других растений (симбиоз), грибами, микроорганизмами,
обитающими в почве (микориза, клубеньки представителей
семейства Бобовые).
8.Синтез биологически
активных веществ.
9.У многих
растений корни выполняют особые функции (воздушные корни, корни-присоски).
Зоны корня — это структуры последовательно сменяющиеся друг друга по мере
роста корня в длину.
Корневой чехлик, или калиптра. Живой наперсток из клеток,
живущих 5-9 дней. Наружные клетки отслаиваются ещё живыми и выделяют обильную
слизь, облегчающую прохождение корня между частицами почвы. На смену им,
изнутри, апикальная меристема продуцирует новые клетки. В клетках осевой части
чехлика, так называемой колумелле, находятся подвижные крахмальные зёрна,
обладающие свойствами кристаллов. Они играют роль статолитов и определяют
геотропические изгибы корней.
Зона деления.
Около 1 мм, прикрыта снаружи чехликом. Она более тёмная или желтоватого
цвета, состоит из мелких многогранных, постоянно делящихся клеток с густой
цитоплазмой и крупным ядром. В зону деления входит апекс корня с его инициалями
и их производными.
Зона роста,
или зона растяжения. Составляет несколько миллиметров, более
светлая, прозрачная. Клетки, пока их клеточные стенки не
станут жёсткими, растягиваются в длину при всасывании воды. Это растяжение
толкает кончик корня дальше в почву. В пределах зоны роста происходит
разделение клеток на ткани.
Зона всасывания, или зона поглощения и дифференциации. До
нескольких сантиметров. Окончание зоны растяжения хорошо заметно по появлению
многочисленных корневых волосков. Хорошо выделяется благодаря развитию
ризодермы, поверхностной ткани, часть клеток которой дает длинные тонкие
выросты — корневые волоски. Они поглощают почвенные растворы в течение
нескольких дней, ниже их формируются новые волоски. Все вместе они создают
впечатление белого пушка вокруг корня. У растения, только что вынутого из
почвы, всегда можно увидеть прилипшие к корневым волоскам комочки почвы. Они
содержат слой протоплазмы, ядро, крупную вакуоль; их тонкие, легко проницаемые
для воды оболочки плотно склеиваются с комочками почвы. Корневые волоски
выделяют в почву различные вещества. Длина варьируется у разных видов растений
от 0,06 до 10 мм. С увеличением влажности почвы образование замедляется;
не образуются они и в очень сухой почве. Корневые волоски появляются в виде
небольших сосочков — выростов клеток. По прошествии определённого времени
корневой волосок отмирает. Продолжительность его жизни не превышает 10—20 дней
Выше зоны
всасывания, там, где исчезают корневые волоски, начинается зона проведения.
Зона проведения. Старая ризодерма отмирает и слущивается. Корень при
этом немного утончается, становится покрытым наружным слоем первичной
коры — экзодермой, выполняющим функцию покровной ткани. По этой части
корня вода и растворы минеральных солей, поглощённые корневыми волосками,
транспортируются в вышележащие отделы растения.
Зона
проведения занимает большую часть корня, она выполняет функции транспорта
растворов из корня в стебель. Здесь видны боковые корни.
Переход одной зоны в другую постепенный и условный.
Корни некоторых
растений имеют склонность к метаморфозу.
Видоизменения
корней:
1.Корнеплод —
утолщённый главный корень. В образовании корнеплода участвуют главный корень и
нижняя часть стебля. Большинство корнеплодных растений — двулетние.
Корнеплоды состоят в основном из запасающей основной ткани (репа, морковь, петрушка).
2.Корневые клубни (корневые
шишки) образуются в результате утолщения боковых и придаточных корней. С их
помощью растение цветёт быстрее.
3.Корни-зацепки —
своеобразные придаточные корни. При помощи этих корней растение «приклеивается»
к любой опоре.
4.Ходульные корни — выполняют
роль опоры.
5.Досковидные корни представляют
собой боковые корни, проходящие у самой поверхности почвы или над ней,
образующие треугольные вертикальные выросты, примыкающие к стволу. Характерны
для крупных деревьев тропического дождевого леса.
6.Воздушные корни — боковые
корни, растут в надземной части. Поглощают дождевую воду и кислород из воздуха. Образуются у
многих тропических растений
в условиях недостатка минеральных солей в почве тропического леса.
7.Микориза —
сожительство корней высших растений с гифами грибов. При таком
взаимовыгодном сожительстве, называемом симбиозом, растение
получает от гриба воду с растворёнными в ней питательными веществами, а
гриб — органические вещества. Микориза характерна для корней многих высших растений, особенно
древесных. Грибные гифы, оплетающие толстые одревесневшие корни деревьев и кустарников,
выполняют функции корневых волосков.
8.Бактериальные клубеньки на корнях высших
растений —
сожительство высших растений с азотфиксирующими бактериями —
представляют собой видоизменённые боковые корни, приспособленные к симбиозу с
бактериями. Бактерии проникают через корневые волоски внутрь молодых корней и
вызывают у них образование клубеньков. При таком симбиотическом сожительстве
бактерии переводят азот, содержащийся в воздухе, в
минеральную форму, доступную для растений. А растения, в свою очередь,
предоставляют бактериям особое местообитание, в котором отсутствует конкуренция
с другими видами почвенных бактерий. Бактерии также используют вещества,
находящиеся в корнях высшего растения. Чаще других бактериальные клубеньки
образуются на корнях растений семейства Бобовые. В связи с этой
особенностью семена бобовых богаты белком, а представителей
семейства широко используют в севообороте для
обогащения почвы азотом.
9.Дыхательные корни — у
тропических растений — выполняюфункцию дополнительного дыхания.
10.Корни-подпорки —
придаточные корни некоторых тропических растений, растущие на стволах и ветвях
и дорастающие до земли[2].
Почвенное
питание растений.
Минеральные
соли поглощаются растением с водой из почвы. Минеральные вещества и вода нужны
для образования многих органических веществ. Нормальное развитие растений
возможно только при наличии таких элементов, как азот, сера, калий, магний,
кальций, железо. Каждый из них имеет индивидуальное значение и не может быть
заменен другим. Если в почве не хватает каких-то элементов, то вносятся
органические и минеральные удобрения. Органические удобрения перегнивают, т.е.
минерализуются, и только после этого усваиваются растениями.
Вопросы
на закрепление
1.
Какие признаки характерны и для стебля, и для корня?
2.
Чем корень отличается от стебля?
3.
Чем отличаются различные корневые системы?
4.
В чем отличие строения и функций различных зон корня?
5.
Для чего удобряют почву? Чем органические удобрения отличаются от
минеральных?
6.
Какое строение имеет корень в зоне всасывания?
7.
Для чего у растений формируются видоизмененные корни?
8.
Корни каких растений мы употребляем в пищу?
9.
Как происходит рост корня в длину и толщину?
Лекция
№4 Морфология листа
Новые
понятия и термины: листовая пластинка, черешок,
прилистники, сидячий лист, сложные и простые листья, листорасположение (
очередное, супротивное, мутовчатое), листья цельные и расчлененные, жилки,
жилкование (параллельное, дуговое, пальчатое, перистое), ловчие аппараты,
листопад, отделительный слой, ксерофиты, мезофиты, гигрофиты.
План
1.Определение
листа
2.Функции
листа
3.Внешнее
строение листа
4.Листорасположение
5.Жилкование
листьев
6.Видоизменения
листьев
7.Старение
листа. Листопад
Лист
— плоский дорсивентральный орган, форма и размеры которого способствуют
созданию максимальной фотосинтезирующей поверхности при оптимальных
значениях транспирации . Количество
листьев на растении весьма различно. Считается, например, что одно дерево дуба несет
до 250000 листьев. Плоская форма делает лист бифациальным, т.е. двусторонним, поэтому
можно говорить о верхней и нижней сторонах листа, имея в виду ориентацию этих
сторон по отношению к верхушке побега . Верхнюю
сторону можно также назвать брюшной, или адаксиальной, а нижнюю — спинной, или
абаксиальной. Это связано с положением листового зачатка в почке . Верхняя и нижняя стороны
нередко существенно различаются между собой по анатомическому строению,
характеру жилкования и окраске.
Размеры листьев чаще всего колеблются в пределах от 3 до 10 см, однако
известны гигантские листья некоторых пальм до 15 м длиной. Крупнейшие листья
известной амазонской кувшинки виктории королевской (Victoria regia) достигают
2 м в диаметре. Размеры, форма и степень рассеченности листьев, хотя и являются
наследственными признаками того или иного вида, очень изменчивы и зависят также
от условий обитания его особей.
Взрослый лист обычно расчленен на пластинку или несколько пластинок (у сложных
листьев) и черешок— узкую стеблевидную его часть,
соединяющую пластинку и узел побега.
Самая нижняя часть листа, сочлененная со стеблем, называется основанием листа .
Часто при основании листа заметны разного размера и формы парные боковые
выросты — прилистники.
Пластинка — главнейшая
часть листа, как правило, осуществляющая его основные функции. Редуцируется
пластинка крайне редко, и тогда ее функции принимают либо расширенный
листовидный черешок — филлодий (у австралийских акаций ),
либо крупные листовидные прилистники (у некоторых видов чины ).
Черешок обычно округлый или
сплюснутый в поперечном сечении. Кроме опорной и проводящей функций он,
длительное время сохраняя способность к вставочному росту, может регулировать
положение пластинки , изгибаясь по направлению к свету.
Нередко черешок не развивается, и тогда лист называют сидячим. Лист с черешком
называют черешковым .
Основание листа принимает различную форму. Весьма часто оно суженное либо имеет
вид небольшого утолщения ( листовая подушечка ).
Однако нередко, особенно у злаков и зонтичных ,
оно разрастается и образует замкнутую или незамкнутую трубку, называемую листовым влагалищем .
Листовое влагалище защищает пазушные почки ,
способствует длительному сохранению интеркалярной меристемы стебля и
нередко служит средством дополнительной опоры побега.
В процессе формирования листа прилистники разрастаются раньше пластинки и
играют защитную роль, составляя часть почечных покровов. После развертывания
почек прилистники часто
опадают или подсыхают. Изредка они имеют размеры, сравнимые с размерами
листовой пластинки (особенно у сложных листьев, в частности, у листьев гороха ), и
функционируют как фотосинтезирующие органы.
В семействе гречишных прилистники
в результате срастания образуют так называемый раструб, охватывающий стебель
над узлом в виде короткой пленчатой трубки.
Итак подводим итог , лист — это боковая часть побега. Типичный лист состоит из
листовой пластинки, черешка и основания. Листовая пластинка выполняет основные
функции листа, черешок — ориентирует листовые пластинки в пространстве,
создавая листовую мозаику, т.е. такое размещение листьев на побеге, при котором
они не затеняют друг друга. Основанием лист прикрепляется к стеблю. Если
черешок отсутствует, лист называют сидячим.
Листья могут быть простыми и сложными. Лист, имеющий одну листовую пластинку,
называют простым. Лист называется сложным, когда на одном черешке располагаются
несколько листовых пластинок на собственных черешочках.
Листовые
пластинки бывают:
Цельными — края листовой пластинки не имеют вырезов.
Расчлененными — края листовой пластинки имеют вырезы различной глубины
Функции листа
1.Фотосинтез
— процесс образования органических веществ (глюкозы) из неорганических
(углекислый газ и вода) под действием солнечного света (при участии хлорофилла)
в фотосинтезирующей паренхиме.
2.Транспирация
— испарение воды растением (эпидерма).
3.Газообмен
— при дыхании поглощается кислород, выделяется углекислый газ, при фотосинтезе
поглощается углекислый газ, а выделяется кислород.
Основные
формы сложных листьев:
1.
Тройчатосложные — к верхушке общего черешка прикрепляются три
листовые пластинки (земляника).
2.
Пальчатосложные — к верхушке общего черешка прикрепляется более
трех листовых пластинок (каштан).
3.
Перистосложные — листовые пластинки располагаются по длине общего
черешка (горох посевной, рябина обыкновенная).
Листорасположение
— это определенный порядок размещения листьев на стебле. Различают три основных
типа листорасположения:
1.
очередное -спиральное, при котором к узлу прикрепляется только
один лист (береза, яблоня).
2.
супротивное, при котором к узлу прикрепляется два листа,
расположенных один против другого (клен).
3.
мутовчатое — к узлу прикрепляется три или больше листьев (вороний
глаз).
Порядок расположения жилок в листе называется жилкованием.
Типы
жилкования:
1.
Дуговое (ландыш). В листе образуется несколько крупных жилок,
которые дугообразно изгибаются и проходят по листу, не соединяясь между собой.
2.
Параллельное (злаки). Крупных жилок в листе несколько. Они
проходят от основания в верхушке листа, не соединяясь между собой, параллельно
друг другу и краю линейного листа.
3.
Пальчатое (смородина). Крупных жилок в листе несколько. Они
расходятся веером от основания листовой пластинки, не соединяясь между собой.
4.
Перистое (береза, черемуха). Крупная жилка одна, от нее отходят
мелкие боковые жилки.
Видоизменения листьев
— выработанные в ходе эволюции необратимые изменения формы листьев в результате
приспособления органов растения к условиям среды обитания (т. е. с выполнением
листьями новых функций).
1. Колючки —
одно из наиболее часто распространенных видоизменений; они служат защитой от
поедания животными. При этом лист либо целиком превращается в колючку (кактусы,
молочаи, барбарис, белая акация, верблюжья колючка), либо в колючку
превращается его часть (бодяк, чертополох, падуб).
2. Усики (у
сложных листьев некоторых видов растений) цепляются за опору, вынося весь побег
к свету. При этом в усик могут превращаться либо верхние листочки сложного
листа (горох, вика), либо весь лист целиком, а функцию фотосинтеза выполняют
прилистники (некоторые виды чины).
3. Запасающую функцию выполняют сочные
чешуи луковиц (лук, чеснок), листья алоэ, кочана капусты.
4. Кроющие чешуи почек
защищают нежные зачаточные листья и конус нарастания от неблагоприятных условий
внешней среды.
5. Ловчие аппараты обеспечивают
жизнь насекомоядных растений на болотах в условиях недостатка азота и других
элементов минерального питания. Листья таких растений изменились до
неузнаваемости, превратившись в ловушки (венерина мухоловка), кувшинчики
(непентес). Листья некоторых растений своими блестящими, ярко окрашенными
капельками на волосках привлекают муравьев, мух, комаров, других мелких
насекомых; выделяющийся при этом сок содержит пищеварительные ферменты
(росянка).
Особенности
растений, произрастающих во влажных и сухих местах
Растения
засушливых мест — ксерофиты. Листья у них мелкие, часто видоизменены в колючки
и чешуйки (кактус). Поверхность листьев покрыта восковым налетом или густо
опушена волосками. У алоэ листья мясистые, запасающие воду.
Растения сильно увлажненных местообитаний — гигрофиты. Листья у них крупные,
тонкие, лишены волосков, со слабо развитой кутикулой, устьица постоянно открыты
(рогоз, бегония).
Растения, произрастающие в условиях достаточной влажности — мезофиты. У них все
характеристики усреднены. К ним относится большинство луговых, плодовых,
овощных растений.
Старение листа. Листопад.
В конце лета у листопадных растений или через несколько лет у вечнозеленых
растений в листьях начинаются процессы старения. В стареющем листе замедляются
процессы синтеза веществ, накапливаются некоторые соли, разрушаются органоиды
клетки, хлорофилл. Лист изменяет окраску на желтую, красную и т.д. У
травянистых растений лист отмирает вместе со стеблем. У древесных растений в
стебле около основания листа образуется специальный отделительный слой. Осенью
клетки этого слоя разрушаются, и лист отделяется от стебля. На месте его
прикрепления к стеблю остается листовой рубец, покрытый пробкой.
Листопад
имеет приспособительное значение:
1.Сокращается
испаряющая поверхность растения.
2.Удаляются
из организма вредные вещества.
3.Растение
избегает поломок под тяжестью снега.
Вопросы
на закрепление
1.
Что такое лист, и какие он функции выполняет?
2.
Опишите внешнее строение листа.
3.
Какие листья называют простыми и сложными?
4.
Каково внутреннее строение листа?
5.
По каким признакам можно отличить лист растения, обитающего в
засушливых условиях, от растения, растущего при избытке влаги?
6.
Как изменяется строение листа перед листопадом и как он
происходит? Значение листопада для растений.
7.
Что такое листовая мозаика и какое значение она имеет для
растений?
8.
Листья каких растений человек употребляет в пищу?
Лекция
№5. Морфология цветка
Новые
понятия и термины: цветоножка, цветоложе, околоцветник
(двойной, простой), чашечка, чашелистики, венчик, лепестки, тычиночная нить,
пыльник, андроцей, завязь, столбик, рыльце, гинецей, верхушечные и боковые
цветки; мужские, женские и обоеполые цветки; однодомные и двудомные растения,
самоопыление, перекрестное опыление, соцветие, типы соцветий: кисть, колос,
сережка, початок, зонтик, щиток, головка, корзинка, метелка, сложный зонтик,
сложный колос
План
ответа:
1.Определение
цветка
2.Функции
цветка
3.Строение
цветка
4.Околоцветник
5.Репродуктивные
органы цветка
6.Классификация
цветков по форме, положению на растении, полу
7.Опыление
и типы опыления
8.
Оплодотворение у растений
9.Двойное
оплодотворение у растений
10.Формулы
цветка
11.Соцветие.
Типы соцветий и их значение.
Цветок — генеративный орган покрытосеменных растений. Он представляет собой
видоизмененный в процессе эволюции укороченный побег с ограниченным ростом.
Функции цветка:
Осуществляет все процессы полового размножения — образование спор, гаметофитов,
опыление, оплодотворение, формирование зародыша, семени, плода.
Строение.
Цветок состоит из стеблевой (цветоножка, цветоложе) и листовых частей (чашелистики,
лепестки, тычинки, пестики). Часть побега, соединяющая цветок со стеблем
называется цветоножкой. Если цветоножка укорочена или отсутствует, то цветок
называют сидячим. Верхняя расширенная часть цветоножки, к которой прикрепляются
все части цветка, называется цветоложем.
Чашелистики и лепестки образуют околоцветник. Околоцветник
может состоять из чашечки и венчика. Чашечка — наружный круг околоцветника,
образованный совокупностью чашелистиков обычно небольших размеров, зеленого
цвета. Она выполняет функцию защиты внутренних частей цветка до раскрывания
бутона. Венчик — это внутренняя часть околоцветника — образован совокупностью
лепестков, более крупных размеров, чем чашечка, разнообразной яркой окраски.
Околоцветник бывает двойным — с чашечкой и венчиком (роза, малина, земляника,
гвоздика), простым венчиковидным — без чашечки (тюльпан, лилия), простым
чашечковидным — без лепестков (свекла, крапива, конопля). Существуют цветки, не
имеющие околоцветника (ясень, осока, ива), их называют голыми.
К репродуктивным частям цветка относят тычинки и пестики. Они
выполняют функцию полового размножения. Тычинки — части цветка, образующие
споры и пыльцу. Тычинка состоит из тычиночной нити и пыльника. Функция
тычиночной нити — прикрепление тычинки к цветоложу и вынос пыльника для лучшего
распространения пыльцы. Пыльник расположен на верхушке тычиночной нити. В нем
формируются пыльцевые мешки, и созревает пыльца (пыльцевые зерна). Совокупность
всех тычинок одного цветка называется андроцеем.
Пестик — репродуктивная часть цветка, состоит из завязи, столбика и
рыльца. Внутри завязи находятся один или несколько семязачатков. Совокупность
пестиков одного цветка называется гинецеем.
Классификация цветков
I. По
форме околоцветника
1.
Правильные – актиноморфные. Через цветок можно провести несколько
осей симметрии (розоцветные, крестоцветные).
2.
Неправильные – зигоморфные. Через цветок можно провести только
одну ось симметрии (горох).
3.
Асимметричные. Нельзя провести ни одной оси симметрии.(Орхидные)
II. По
расположению на побеге
1.
Верхушечные — образуются на верхушке побега.
2.
Боковые — располагаются в пазухе листьев по бокам побега.
III.
По половой принадлежности.
1.
Мужские – тычиночные цветки имеют только андроцей.
2.
Женские – пестичные цветки имеют только гинецей.
3.
Обоеполые цветки (андроцей и гинецей в одном цветке).
Если на одном растении располагаются и женские и мужские цветки, оно называется
однодомным (кукуруза, огурец).
Если
мужские цветки располагаются на одном растении, а женские на другом растение
называется двудомным (тополь, ива, облепиха).
Для краткой записи строения цветка используются специальные формулы, в
которых указываются части цветка, их количество и особенности расположения и
строения.
Чашелистики обозначаются буквой Ч, лепестки венчика — Л, тычинки — Т, пестики —
П. Их количество — цифрой около соответствующей буквы, знак бесконечности
проставляется в том случае, если частей цветка больше 10. Срастание частей
цветка — скобками, знаком + — расположение частей цветка в разных кругах,
запятая ставится между частями, отличающимися своим строением. Перед формулой
правильных цветков ставится знак *, а неправильных — стрелка.
Процесс переноса пыльцы на рыльце пестика называется опылением.
При самоопылении на пестик попадает пыльца того же растения. Если перенос
пыльцы осуществляется между цветками разных особей, то происходит перекрестное
опыление. Оно происходит у 90% растений, т.к. повышает разнообразие потомков.
Поэтому в процессе эволюции выработался ряд приспособлений, ограничивающих
самоопыление:
1.
двудомность.
2.
созревание пыльцы и семязачатка в разное время.
3.
расположение рыльца пестика и пыльника на разной высоте.
4.
неспособность пыльцы прорастать на рыльце пестика своего цветка.
При эволюции цветковых растений все
большую и большую роль в перенесении пыльцы играли насекомые. Необходимость
привлечения насекомых (и мелких птиц) как эффективных переносчиков пыльцы
является одной из причин образования цветка.
Двойное
оплодотворение у растений
Уникальная особенность цветковых растений — двойное оплодотворение.
В завязь покрытосеменных растений проникает два спермия, один из них сливается
с яйцеклеткой, дав начало диплоидному зародышу. Другой соединяется с
центральной диплоидной клеткой. Образуется триплоидная клетка, из которой
возникнет эндосперм — питательный материал для
развивающегося зародыш. Этот процесс, характерный для всех покрытосеменных,
открыт в конце прошлого века С.Г. Навашиным и получил название двойного оплодотворения.
Значение двойного оплодотворения, по- видимому, заключается в том, что
обеспечивается активное развитие питательной ткани уже после оплодотворения.
Поэтому семяпочка у покрытосеменных не запасает питательных веществ впрок и,
следовательно, развивается гораздо быстрее, чем у многих других растений,
например у голосеменных.
У цветковых растений имеется ряд особенностей образования половых клеток и
оплодотворения. Оплодотворению у них предшествует образование сильно
редуцированного гаплоидного поколения — гаметофитов.
После оплодотворения прорастание пыльцы цветковых растений начинается с разбухания зерна и
образования пыльцевой трубки , которая прорывает спородерму в
более тонком ее месте — так называемой апертуре. Кончик пыльцевой трубки выделяет
специальные вещества, размягчающие ткани рыльца и столбика , в которые внедряется пыльцевая
трубка. По мере роста пыльцевой трубки в нее переходят ядро вегетативной клетки
и оба спермия . В огромном большинстве
случаев пыльцевая трубка проникает в мегаспорангий ( нуцеллус )
через микропиле семязачатка , реже —
иным образом. Проникнув в зародышевый мешок , пыльцевая трубка разрывается, и ее
содержимое изливается внутрь. Один изспермиев сливается с яйцеклеткой, и
образуется диплоидная зигота, дающая затем начало зародышу . Второй
спермий сливается со вторичным ядром, располагающимся в центре зародышевого мешка ,
что приводит к образованию триплоидного ядра, развивающегося затем в
триплоидный эндосперм. Весь этот процесс получил название двойного
оплодотворения. Он был впервые описан в 1898 г. выдающимся русским цитологом и
эмбриологом С.Г.Навашиным. Прочие клетки зародышевого мешка — антиподы и синергиды в
оплодотворении не участвуют и довольно быстро разрушаются.
Как и у других высших растений , при мейозе у цветковых
образуются споры . Микроспоры (мужские
споры) образуются в пыльниках , из них
формируются пыльцевые зерна (мужские
гаметофиты). Зрелое пыльцевое зерно состоит из трех клеток — вегетативной (или
клетки, пыльцевой трубки ) и двух спермиев(лишенные жгутика
сперматозоиды). Мегаспоры (женские
споры) формируются в семяпочках внутри завязи пестика .
Биологический смысл двойного оплодотворения весьма велик. В отличие от голосеменных , где
довольно мощный гаплоидный эндосперм развивается независимо от процесса
оплодотворения, у покрытосеменных триплоидный эндосперм образуется лишь в
случае оплодотворения. С учетом гигантского числа поколений этим достигается
существенная экономия энергетических ресурсов. Увеличение же уровня плоидности
эндосперма до 3n, по-видимому, способствует более быстрому росту этой
полиплоидной ткани по сравнению с диплоидными тканями спорофита .
Взаимодействие пыльцевой трубки гаметофита с тканями спорофита — сложный процесс,
регулируемый химическими веществами. Так, выяснилось, что если промыть пыльцу
дистиллированной водой, она теряет способность к прорастанию. Если
сконцентрировать полученный раствор и обработать концентратом пыльцу, она вновь
станет полноценной. После прорастания рост пыльцевой трубки контролируется
тканями пестика. Например, у хлопчатника рост
трубки до яйцеклетки занимает 12-18 ч, но уже через 6 ч можно установить, к
какой семяпочке направляется пыльцевая трубка: в этой семяпочке начинается
разрушение особой клетки — синергиды . Как
растение направляет рост трубки в нужную сторону и каким образом синергида
узнает о ее приближении, пока еще не известно.
Во многих случаях у цветковых растений существует «запрет» на
самоопыление: спорофит «узнает» своего мужского гаметофита и не
разрешает ему участвовать в оплодотворении. В некоторых случаях при этом
собственная пыльца не прорастает на рыльце пестика. В большинстве же случаев
рост пыльцевой трубки начинается, но затем останавливается и она не достигает
яйцеклетки. Например, у первоцвета весеннего
еще Ч.Дарвин обнаружил две формы цветков — длинностолбиковые (с длинным
столбиком и короткими тычинками) и короткостолбиковые (столбик короткий,
тычиночные нити длинные). У короткостолбиковых растений пыльца почти вдвое
крупнее, а клетки сосочков рыльца мелкие. Все эти признаки контролируются
группой тесно сцепленных генов.
Опыление эффективно только при переносе пыльцы с одной формы на другую. За
распознавание своей пыльцы отвечают молекулы-рецепторы, представляющие собой
сложные комплексы белков с углеводами. Показано, что растения дикой капусты ,
которые не вырабатывают в тканях рыльца молекул рецептора, могут самоопыляться.
У нормальных растений рецепторы появляются на рыльце за день до открытия
цветка. Если раскрыть бутон и нанести на него собственную пыльцу за два дня до
распускания, то оплодотворение произойдет, а если за один день до распускания —
то нет.
Интересно, что в некоторых случаях самонесовместимость пыльцы у растений
определяется серией множественных аллелей одного гена, сходно с
несовместимостью при пересадках тканей у животных. Эти аллели обозначаются
буквой S, и число их в популяции может достигать десятков и даже сотен. Если,
например, генотип производящего яйцеклетки растения — s1s2, а производящего
пыльцу — s2s3, то прорастать при перекрестном опылении будут только 50% пылинок
— те, что несут аллель s3. При наличии десятков аллелей большая часть пыльцы
при перекрестном опылении нормально прорастает, а самоопыление полностью
предотвращается.
Соцветия, их типы и биологическая роль
Соцветия — это группа из нескольких цветков, расположенных на одном цветоносном
стебле.
Типы соцветий:
Кисть — соцветие, у которого на длинной оси располагаются цветки на цветоножках
одинаковой длины (черемуха).
Колос — на длинной оси колоса располагаются сидячие цветки (подорожник).
Початок — Вытянутая ось соцветия утолщена, и на ней располагаются сидячие
цветки (кукуруза).
Зонтик — цветки имеют цветоножки одинаковой длины, которые прикрепляются на
верхушке главной оси (лук).
Щиток — на длинной оси соцветия располагаются цветки, у которых цветоножки
имеют различную длину, поэтому все цветки расположены в одной плоскости
(рябина).
Головка — соцветие имеют короткую, утолщенную ось, а на ней расположены сидячие
цветки (клевер).
Корзинка — ось соцветия имеют форму блюдца, на ней располагаются сидячие цветки
(подсолнечник).
Сложный зонтик — к верхушке оси соцветия прикрепляются не цветки, а зонтики
(укроп).
Сложный колос — на оси колоса располагаются не цветки, а колосья (пшеница).
Сережка — это повислый колос (береза).
Метелка — соцветие, у которого на оси располагаются кисти или щитки (овес,
сирень).
Значение соцветий:
Цветки
в соцветии заметнее для насекомых опылителей.
Увеличение
вероятности опыления ветром.
Последовательное
распускание цветков имеет преимущества: если при цветении первых цветков
условия для опыления были неблагоприятными, то последующие могут
опылиться.
Вопросы
на закрепление:
1.
Что такое цветок, какие функции он выполняет?
2.
Из каких частей состоит околоцветник?
3.
Какие части цветка называют репродуктивными? Каково их строение?
4.
Разделите цветки на группы по расположению на растении, по форме,
по полу.
5.
В чем отличие опыления от оплодотворения? Какие типы опыления вам известны?
6.
В чем заключается биологический смысл двойного оплодотворения у
растений?
7.
Какие приспособления выработались у растений, препятствующие
самоопылению?
8.
Приведите примеры растений с различными типами соцветий. Каково
значение соцветий?
9.
Какое растение имеет самый большой цветок?
Лекция
№6. Плод
Новые
понятия и термины: плод, околоплодник, плодоножка,
листовка, орешек, многоорешек, боб, костянка, многокостянка. орех, коробочка,
стручок, стручочек, зерновка, семянка, ягода, тыквина, яблоко, односемянные и
многосемянные плоды, сухие и сочные плоды, вскрывающиеся и невскрывающиеся
плоды
План
ответа:
1.Определение
плода
2.Функции
плода
3.Образование
4.Строение
плода
5.Классификация
плодов
6.Распространение
плодов и семян
Плод — это орган размножения и расселения покрытосеменных растений,
развивающийся из цветка после оплодотворения.
Функции
плодов: защита формирующихся и распространение зрелых семян.
Плод образуется из завязи пестика. Стенки завязи видоизменяются и формируют околоплодник,
окружающий семена. Цветоножка превращается в плодоножку. У многих растений в
образовании плода участвуют цветоложе, основания тычинок, лепестков,
чашелистиков.
Строение.
Плод
состоит из семян, окруженных околоплодником.
Краткая характеристика различных типов плодов:
Орешек — сухой односемянный плод, невскрывающийся. Околоплодник твердый
деревянистый. (береза, гречиха) Особый вид многоорешка — плод земляники, у
которого орешки расположены на поверхности мясистого цветоложа. У шиповника
многоорешки располагаются внутри мясистого цветоложа.
Боб — сухой многосемянный плод, свойственный бобовым. При созревании
раскрывается по двум швам (брюшному и спинному), распадаясь на две части.
Семена прикрепляются к створкам (горох). Бобы могут не вскрываться створками, а
разделяться на части, содержащие по одному семени.
Костянка — сочный односемянный плод, невскрывающийся. Для нее характерна
твердая косточка, образовавшаяся из внутренних слоев околоплодника (слива,
вишня, персик, черемуха). Плод, состоящий из многих костянок, расположенных на
общем цветоложе, называют многокостянкой (малина, еже¬вика, костяника).
Орех — крупный односемянный плод с деревянистым твердым околоплодником (фундук,
лещина).
Коробочка — многосемянный плод с сухим вскрывающимся околоплодником. (зверобой,
табак, мак, белена, подорожник).
Стручок — сухой многосемянный плод. Состоит из двух створок и центральной
перегородки, на которой расположены семена. Вскрывается снизу вверх, разделяясь
на две части, между которыми располагается перегородка с семенами
(крестоцветные).
Стручочки — это стручки, у которых длина приблизительно равна ширине (пастушья
сумка).
Зерновка — односемянный плод, невскрывающийся, у которого тонкий пленчатый
околоплодник настолько тесно прилегает к семенной кожуре, что кажется сросшимся
с ней (злаки).
Семянка — односемянный плод с кожистым околоплодником, легко отделяющимся от
семени. Он содержит одно семя, не вскрывается при созревании (сложноцветные).
Ягода — многосемянный плод, в зрелом состоянии имеет мясистый сочный
околоплодник (виноград, томат, паслен, картофель, брусника, голубика, клюква).
Тыквина — плод, близкий к ягоде, но отличающийся более плотным околоплодником
(тыква, арбуз, огурец).
Яблоко — многосемянный, невскрывающийся плод, с более плотным, чем у ягоды,
околоплодником (яблоня, рябина, боярышник).
Плоды
различаются по следующим признакам:
1. По
консистенции околоплодника:
A.
Сухие плоды (листовка, орешек, стручок, боб).
B. Сочные плоды (ягода, костянка, тыквина, яблоко).
2. По
количеству семян в плоде:
A.
Односемянные (семянка, орех, зерновка).
B. Многосемянные (ягоды, яблоко, стручок).
3. По
способу вскрывания:
A.
Невскрывающиеся (орешек, семянка).
B. Вскрывающиеся (коробочка, стручок, боб).
Соплодие
— это совокупность сросшихся между собой зрелых плодов, которые выглядят как
один плод (ананас, свекла, инжир, шелковица).
Распространение плодов и семян
1.плоды
и семена распространяются ветром. У них обычно образуются различные крылатые
выросты — летучки (плоды одуванчика, клена, березы; семена тополя, осины, ивы).
2.распространение
с помощью воды. Семена и плоды имеют выросты, заполненные воздухом или волоски,
не смачивающиеся водой (кувшинка ивы).
3.распространение
с помощью животных. Эти плоды и семена могут поедаться животными (сочные плоды)
или имеют приспособления для прикрепления к покровам животных (прицепки у
лопуха, череды, клейкие семена подорожника).
4.саморазбрасывание
семян у недотроги, фиалок, кислицы, бешенного огурца.
Вопросы
на закрепление:
1.
Что такое плод? Какие функции он выполняет?
2.
Из чего образуется плод?
3.
Назовите все части плода и их функции.
4.
Охарактеризуйте сухие и сочные плоды.
5.
Сравните вскрывающиеся и невскрывающиеся плоды.
6.
Каким образом могут распространяться плоды и семена.
7.
Какое растение имеет самый крупный плод?
8.
Плоды каких растений человек употребляет в пищу? К каким типам
плодов они относятся?
Лекция
№7. Семя
Новые
понятия и термины: микропиле, двудольные и однодольные
растения, семядоля; зародышевый корешок, стебелек и почечка; щиток, всхожесть и
покой ,партенокарпия, партеногенез, партеноспермия
План
ответа:
1.Определение
семени.
2.Происхождение
частей семени.
3.Строение
семени.
4.Отличие
семян Однодольных от семян Двудольных растений.
5.Состояние
покоя. Прорастание семян
6.
Партенокарпия, партеногенез, партеноспермия
СЕМЯ
(semen) — орган голосеменных и цветковых растений, образующийся из семяпочки
и выполняющий функции воспроизведения, расселения и переживания неблагоприятных
условий. У большинства растений условие образования семени — оплодотворение
(иногда семя образуются без оплодотворения, в результате апомиксиса). У
цветковых растений семяпочки находятся в завязях; у голосеменных семяпочки
располагаются открыто на поверхности семенных чешуек шишки (мегастробила).
Обычно семя состоит из семенной кожуры, зародыша и у многих растений из тканей
с запасными питательными веществами — эндосперма или перисперма (часто
питательные вещества откладываются в семядолях зародыша). У зрелого семени
зародыш представляет собой миниатюрное растение и состоит из зародышевого
корешка, зародышевого стебелька (гипокотиля), семядолей (зародышевых листьев,
нередко с запасающей функцией) и почечки, в которой различают конус нарастания
и у многих растений зачаточные листья.
По числу семядолей зародыша цветковые растения
делятся на однодольные и двудольные , в зародыше голосеменных растений от 2 до
18 семядолей. Эндосперм у голосеменных — вегетативная часть женского заростка с
гаплоидным набором хромосом, у цветковых — триплоидное новообразование,
возникающее в результате слияния второго спермия с вторичным ядром зародышевого
мешка. У одних видов в ткани эндосперма преобладают углеводы и белки ( у дуба),
у других — белки и жиры (у сосны, ореха ). Кожура семени защищающая зародыш от
высыхания и поражения вредителями и болезнями, может быть тонкой и мягкой (у
тополя, ивы ) или твёрдой и толстой ( у гледичии, боярышника). Она иногда имеет
выросты в виде шипов, волосков и крылышек, способствующих рассеиванию семян.
Таким образом , семя покрытосеменных растений — это
видоизмененный в процессе двойного оплодотворения семязачаток.
Строение
семени и происхождение его частей. Семя состоит из
зародыша и запасных питательных веществ, покрытых семенной кожурой. Зародыш
развивается из оплодотворенной яйцеклетки, т.е. из зиготы. Из оплодотворенного
центрального ядра формируется эндосперм. Покровы семяпочки превращаются в
кожуру семени.
Семенная кожура. Покрывает семя снаружи, обеспечивая защиту. Если на ней
образуются выросты, то семенная кожура участвует в расселении семян.
Запасающие питательные ткани содержат масла (у масличных растений), крахмал (у
злаков) или белки (у бобовых). Как правило, запасающей питательной тканью у
растений является эндосперм. Эндосперм чаще твердый.
Зародыш — это будущее растение. Он состоит из зародышевого корешка, стебелька и
почечки. Первые листья зародыша называются семядоли.
Строение семян двудольных и однодольных растений.
Главное отличие семян заключается в строении зародыша.
Двудольные растения. Зародыш у них имеет две семядоли, между которыми
находится конус нарастания зародышевого стебля. Семядоли покрыты эпидермисом и
заполнены клетками, богатыми запасными питательными веществами, которые
используются растением при прорастании семян.
Однодольные растения. Зародыш имеет одну семядолю. У злаков единственная
видоизмененная семядоля зародыша называется щиток. Щиток прилегает к эндосперму
Прорастание семян
при наличии в окружающей среде достаточного
кол-ва воды и тепла. В лесу прорастает не более 1 — 2% выпавших семян, а при
посеве в лесопитомниках, где создаются благоприятные условия, может прорасти
40—80% С. Различают надземное прорастание, когда гипокотиль выносит семядоли
над почвой (напр., у сосны, ели, липы, клёна) и подземное, при котором семядоли
остаются под землёй, а на поверхность выходит побег (его первое междоузлие
называется эпикотилем), растущий из почки (напр., у дуба).
Семя ношение у древесных пород начинается в
возрасте 10—15 лет и продолжается до старости.
Жизнеспособность семян в естественных условиях
сохраняется от 0,5—1,5 мес (у тополя) до 2—3 лет (у дуба, лещины и др.), однако
большая часть семян, выпадающих осенью и зимой, прорастает весной или в
первой половине вегетационного периода, следующего за годом их созревания.
Качество семян. определяется массой 1 тыс. штук, энергией прорастания,
лабораторной и грунтовой всхожестью, а также чистотой. У древесных пород перед
посевом в питомниках семена подвергаются стратификации.
Далеко не все семена обладают способностью прорастать сразу после отделения от
материнского растения. У большинства цветковых растений созревшие семена
переходят в состояние покоя. Это очень важно в условиях сезонного климата,
чтобы проростки не погибали в неблагоприятных условиях осенью и зимой.
Выявлено несколько причин, по которым семена не прорастают
1.Недоразвитие
зародыша
2.Накопление
веществ, затормаживающих рост.
3.Очень
плотные оболочки, защищающие от попадания влаги.
Многие семена сохраняют всхожесть (способность к прорастанию) до нескольких
десятков лет, создавая запас семян в почве.
Прорастание обычно начинается с набухания семени и появления корня из-под
покровов семени, затем начинают расти и зеленеть семядоли, из зародышевого
стебелька развивается побег.
Как только в семя поступает вода, в нем резко усиливается дыхание, и
активизируются вещества, которые превращают запасные питательные вещества в
усваиваемую клеткой форму. Питательные вещества перемещаются в зародыш и
используются для деления, роста и жизнедеятельности клеток.
Условия прорастания семян:
1.Влажность.
Для активизации ферментов, ускоряющих поглощение запасных питательных веществ
зародышем.
2.Воздух
(кислород) для перехода питательных веществ в усвояемую форму.
3.Солнечный
свет нужен не для всех растений.
4.Тепло.
Для различных видов существует свой температурный оптимум.
В лесах семена растений распространяются с помощью
птиц и животных (зоохория) насекомых (энтохория), ветра (анемохория) и воды
(гидрохория).
Особую форму полового
размножения представляет собой партеногенез, или девственное размножение, —
развитие организма из неоплодотворенной яйцеклетки.
Данная форма размножения характерна
главным образом для видов, обладающих коротким жизненным циклом с выраженными
сезонными изменениями.
Партеногенез бывает гаплоидным и диплоидным.
В природе естественный
партеногенез встречается у ряда растений (одуванчик, ястребинка и
др.) и носит название апомиксис Под апомиксисом обычно понимают либо развитие
из неоплодотворенной яйцеклетки, либо возникновение зародыша вообще не
из гамет (например,
у цветковых растений зародыш может развиваться
из различных клеток зародышевого мешка)-
Партенокарпия (от греч. parthenos — «девственница»
и karpos — плод; дословно — «девственный плод») —
частный случай партеногенеза,
девственное оплодотворение без
опыления у растений, обычно
с образованием плодов без семян. Партенокарпия
является широко распространеным явлением у плодовых, цитрусовых и овощных
растений.
Партенокарпия, как и большинство типов стерильности,
явление регрессивное и не имеет эволюционного значения.
Различают вегетативную
партенокарпию, также называемую автономной, когда плоды на растении
завязываются и в дальнейшем развиваются без непосредственного процесса опыления. Также выделят стимулятивную
партенокарпию, при которой для образования плодов необходимо раздражение рыльца
цветков пыльцой от других видов растений.
К примеру, пыльца яблони способна
вызвать партенокарпию у груши, а
пыльца томата —
у баклажана.
Данный тип партенокарпии может быть вызван искусственно путём механических,
химических, либо тепловых раздражений. Благодаря этому она имеет определенное
хозяйственное значение, так как получаемые таким способ плоды характеризуются
своим обилием, большой сочностью, мясистостью и высокими вкусовыми качествами,
а также имеют преимущества при технологической обработке.
Плоды, которые образуются при партенокарпии, либо бессемянные или содержат
«пустые» семена без зародышей. Растения, характеризующиеся развитием только
бессемянных плодов, размножаются исключительно вегетативным путём.
Партенокарпия известна у многих культурных растений и
часто является прочно закреплённым сортовым признаком. Встречается у ряда
сортов винограда,
яблони, груши, томатов, мандаринов, апельсинов,
банана, крыжовника, хурмы, огурца и
многих друг
Партеноспермия — у хвойных, особенно у лиственницы, при отсутствии опыления и
оплодотворения иногда формируются семена, не содержащие зародыша , что
отрицательно влияет на качество семян.
Вопросы
на закрепление
1.
Что называют семенем, из каких частей оно состоит?
2.
Из чего образуются и чем отличаются семена голосеменных и
покрытосеменных растений?
3.
Чем отличаются семена однодольных и двудольных растений?
4.
Какое биологическое значение имеет состояние покоя у семян?
5.
Какие условия необходимы для прорастания семян?
6.
У каких растений впервые появилось семя?
7.
Семена каких растений использует человек?
8.
Семя мари имеет форму колеса, а марьянника — куколки муравья.
Почему могла возникнуть такая форма семян?
Лекция
№8. Способы размножения растений. Семенное
размножение
Новые
понятия и термины: гамета, зигота, регенерация,
клон, стеблевые отводки, выводковые почки,отпрыски,прививка,подвой,привой,
План
1.Введение
2.Типы
размножения растений
3.Семенное
размножение
4.Вегетативное
размножение
5.Прививки
6.
Значение размножения в природе
При
семенном размножении не всегда передаются признаки сорта, например, махровость
и пестролистность. Недостатком семенного размножения является также более
позднее цветение. Однако оно дает возможность получить новые сорта. Семенами
размножают немногие растения (кактусы, пальмы, лавр, кофе и др.). Семена для
размножения необходимо собирать со здоровых растений с красивыми цветами,
обладающих всеми теми качествами, которые вы хотели бы иметь у будущих
выращенных культур. Плоды с семенами собирают после их вызревания в теплое и
сухое время года и кладут в матерчатый или бумажный мешочек. Спустя несколько
дней семена извлекают из плодов и хранят в бумажном пакете в прохладном сухом
месте (можно в холодильнике).
Чтобы
убедиться, годны ли семена к севу, их необходимо опустить в стакан с водой.
Семена, потерявшие способность к прорастанию, останутся на поверхности воды, а
способные прорасти опустятся на дно.
Перед
посевом семена с твердой оболочкой, например, у пальм и акаций, слегка
подпиливают, чтобы обеспечить доступ воды и воздуха к зародышу и ускорить
прорастание. Семена некоторых субтропических растений (камелии, фейхоа, чая)
нуждаются в стратификации. Их помещают в плошку с влажным песком, все
заворачивают в полиэтиленовую пленку и хранят в холодильнике при температуре
6—8°С около двух месяцев, периодически увлажняя.
В
комнатах можно выращивать из семян амариллис, бегонию клубневую, бальзамин,
кливию, цикламен, глоксинию, кофейное деревце, пеларгонию и многие другие
растения. Все названные виды в условиях домашнего содержания дают всхожие
семена. Их высевают в маленькие горшочки, плошки и деревянные ящики с
водосточными отверстиями. Каждое отверстие прикрывают черепком, а в горшках
делают дренаж высотой не менее 1—2 см из черепков, камешков или угольков. Этот слой
предохраняет водосточные отверстия от засорения землей и способствует, таким
образом, удалению излишней влаги. Посуду наполняют земляной смесью (2 части
листовой земли, 1 часть легкой дерновой и 1/2 части речного песка), поверхность
хорошо выравнивают, слегка уплотняют и приступают к посеву. Земля должна быть
без грубых комков, но просеивать ее через мелкие сита не рекомендуется, так как
в этом случае она быстро закисает. Легче всего сеять крупные семена: их не
слишком густо размещают на равных друг от друга расстояниях. Твердокожие семена
перед посевом кладут на 4—6 дней в теплую воду, кожица набухает, и появление
ростков облегчается.
После
посева семена средней величины засыпают слоем земли, толщина которого равна или
несколько больше толщины семени. Затем землю выравнивают гладкой дощечкой. Чем
тяжелее земля, тем слой покрытия должен быть тоньше. Однако при слишком тонком
покрытии корешок приподнимает семя и выталкивает его из земли, вследствие чего
молодое растение может погибнуть. Хорошо заделать семена просеянной через
мелкую сетку торфяной землей. Она пористая, рыхлая и дает нежным всходам
большую устойчивость влаги. Крупные семена можно погрузить в землю на 2—4 см
(пальмы, лавр и др.). Мелкие (бегония, глоксиния, пеперомия, мамиллярия и др.)
принято не засыпать землей, так как при поливе они сами достаточно углубляются
в почву.
Для
некоторых семян, например, бегонии клубневой, землю предварительно нужно
прокалить, чтобы устранить развитие плесени и мха, которые препятствуют
нормальному развитию молодых сеянцев.
Засыпанные
землей крупные семена поливают теплой водой из лейки с мелким ситом, не
засыпанные мелкие — с большой осторожностью увлажняют из пульверизатора (лучше
наливать воду в поддоны). Посевы накрывают стеклом и поддерживают равномерную
влажность.
Емкости
с высеянными семенами ставят в теплое место в тени, где температура мало
изменяется. Можно заменить стекло слоем влажного мха, который сохранит для
посевов хорошую аэрацию (доступ воздуха). Нужно внимательно следить за
прорастанием семян, чтобы вовремя поднять мох (или снять стекло) во избежание
вытягивания проростков.
Весной
горшки ставят в удаленное от света место с целью сохранения влаги. Если земля
слишком сухая, всходы не развиваются, если слишком влажная — начинают загнивать.
Поливают проростки лейкой с мелким ситечком, а лучше всего погрузить горшок в
емкость с водой, но при этом следить, чтобы уровень воды в ней был ниже
верхнего края горшка на 2 см. Этот способ полива позволит тщательно пропитать
весь земляной ком, и выгоден тем, что ростки избегают механического воздействия
воды.
Семена
большинства видов содержат запас питательных и других веществ, поэтому в первый
период роста сеянец в питательных веществах из внешней среды не нуждается. Как
известно, для нормального развития растению необходимы вода, доступ воздуха,
нужная температура. Температура, при которой удачно прорастают семена различных
растений, тесно связана с географическим происхождением (родиной) данного вида.
Семена пальм и других представителей тропиков прорастают при 22—26°С. Долго и
трудно прорастающие семена следует сеять в январе и феврале, остальные — в
марте и апреле. Слишком поздний посев не рекомендуется, так как в этом случае
сеянцы не успевают достаточно развиться до наступления осени.
У
большинства комнатных растений всходы появляются через 2—3 недели, у некоторых
— через несколько дней, у пальм — через несколько месяцев. С появлением всходов
стеклянное укрытие снимают, и нежные растеньица постепенно приучают к свету.
Температуру после прорастания семян снижают на 3—5°С, чтобы не допустить
вытягивания молодых сеянцев. Более крупные сеянцы можно высадить в горшочки
диаметром 7 см, а мелкие распикировать (рассадить) на расстояние, зависящее от
величины и быстроты развития данного растения. Мелкие всходы, например, бегоний
и глоксиний, нужно пикировать несколько раз, пока они не разовьются, затем
посадить поодиночке в горшки. Рассаживают мелкие сеянцы с помощью пикировочного
колышка и пинцета.
Укорачивание
(прищипка) корешков у сеянцев несколько задерживает их рост, но зато со
временем корни разрастаются в разветвленную мочковатую корневую систему,
которая обеспечивает прекрасное развитие надземной части растений и их обильное
цветение. По мере роста сеянцев их иногда по нескольку раз переваливают из одного
горшка в другой большего размера и обязательно подкармливают. Некоторым молодым
растениям необходимо довольно частое опрыскивание.
Вегетативное размножение растений
Размножение отводками
Размножение древесных
растений отводками — один из самых старых способов, используемых в садоводстве.
Он заключается в стимулировании образования корней на стебле до того, как его
отделят от материнского растения. Метод достаточно прост, поскольку при нем нет
необходимости тщательно контролировать условия окружающей среды, чтобы
поддержать жизнеспособность побегов во время формирования корней.
Как и при любом другом способе размножения, для
отводков важно правильно подобрать побеги, а также участок с соответствующими
почвенными условиями, где они смогли бы успешно укорениться. Лучшему
корнеобразованию способствует и тепло, поэтому отводки целесообразно
располагать на участке, куда проникает достаточно солнечных лучей.
Обычные отводки
Закладка обычных отводков — самый простой и
эффективный метод размножения большого числа древесных пород. Этот метод связан
с минимальным повреждением родительского растения.
Ветку засыпают землей, оставляя лишь верхушку.
Там, где побег присыпан землей, и образуются корни. Как только сформируется
достаточное количество корней, отводок можно отделять от маточного растения.
Развитие корней на подземной части стебля связано с усиленным притоком туда
питательных веществ и фитогормонов. Чтобы успех был гарантирован, нужно
использовать побеги с развитой способностью к корнеобразованию и располагать их
неглубоко под поверхностью земли.
За двенадцать месяцев до закладки отводков, а
иногда и раньше проводят сильную обрезку ветвей. Это способствует быстрому
отрастанию молодых побегов — они лучше всего подходят для образования отводков,
так как их легче пригибать к земле; кроме того, они быстрее окореняются.
Отводки обычно закладывают в начале весны, как только почву станет можно
обрабатывать.
Поскольку размножение отводками обычно проводят
не в специальном питомнике, а просто в саду, важно очень тщательно приготовить
почву. Перекапывают ее как можно глубже. Затем вносят в нужных количествах торф
и песок, превращая тем самым обычную почву в среду с оптимальными условиями для
корнеобразования: хорошей водоудерживающей способностью, воздушным режимом и
дренажем.
На отрезке побега в 10—60 см, считая от верхушки,
удаляют все листья и боковые побеги. Ветку пригибают и отмечают на земле ее
положение примерно в 20 см от ее верхушки. От указанной точки выкапывают
канавку глубиной вначале 10—15 см и по мере приближения к маточному растению
постепенно сходящую на нет.
Секрет образования корней заключается в нарушении
перемещения в стебле питательных веществ и фитогормонов. Для этого обычно
бывает достаточно согнуть стебель под прямым углом. Однако у трудноокореняемых
культур сначала прибегают к кольцеванию стебля ножом или обвязыванию побега в
месте сгиба медной проволокой.
Ветку укладывают на дно выкопанной канавки,
оставляя снаружи загнутый под прямым углом верхний конец длиною около 20 см. Если
стебель «непослушен» и выгибается назад, его закрепляют проволочными дужками.
Сверху засыпают землей, слегка утрамбовывают и поливают.
За влажностью почвы нужно следить, особенно в
засушливые периоды. Окоренение отводков обычно происходит в период вегетации.
Осенью отводок отделяют от маточного растения, чтобы вновь образовавшееся
растение приспособилось к условиям «независимого» существования.
Тремя-четырьмя неделями позже для лучшего
развития корней верхушку отводка обрезают. Затем отводок пересаживают в горшок
или на новое место. Если же к осени корневая система отводка еще достаточно не
развилась, его не выкапывают до следующего года.
Воздушные отводки
Воздушные отводки —
очень интересный способ приумножить количество растений в своей коллекции. Размножение
воздушными отводками необычный способ применимый для многих растений.
Часто он используется в тех случаях, когда длина растения становится
избыточной.
Стебли и ветки некоторых
кустарников и деревьев не удается пригнуть к земле. Их можно размножить,
формируя отводки в воздухе. Подготовьте молодой побег, как простой отводок, но
держите открытым разрез, подсунув в него комочек влажного мха-сфагнума (его
можно приобрести в цветочном магазине). Возьмите больший клочок мха и оберните
им подготовленную часть стебля, закрепив полиэтиленовой пленкой. Когда появятся
корни (через один-два года), постарайтесь это не пропустить. Обработайте корни,
как уже было описано. Снимите пленку, а мох оставьте, чтобы не повредить нежные
корешки.
Горизонтальные
отводки
Горизонтальные отводки представляют
собой побеги растения, уложенные на землю и прикрепленные к ней в 3-5 местах
шпильками из проволоки по всей длине. Сверху побеги присыпаются землей. После
укоренения так же отделяются и пересаживаются. Хорошо размножаются розы,
гортензии и др.

Дугообразные
отводки
Дугообразные отводки — разновидность
горизонтальных. Разница лишь в том, что побеги прикрепляются к земле в одном
месте, ближе к верхнему краю, образуя стеблем над землей дугу.

Размножение черенками
Выращивание растений из
стеблевых черенков — самый популярный способ вегетативного размножения. Однако
его распространение приходится лишь на последние 150 лет благодаря появлению
доступных и дешевых материалов, развитию тепличного хозяйства, парников и т. д.
Основная трудность при
размножении стеблевыми черенками — это поддержание жизнеспособности отделенного
от маточного растения черенка до тех пор, пока он окоренится и превратится в
новое растение. В этом и заключается главное отличие черенкования от метода
размножения отводками, при котором стебель не отделяют от материнского
растения, пока у него не образуется собственная корневая система.
Материал для черенкования
Способность стебля
образовывать корни зависит от возраста материнского растения и сорта, а также
от типа побега. Садовод скорее станет размножать недавно сформировавшееся
растение, являющееся сравнительно новым сортом, чем растение старое или старый
сорт.
Маточное растение коротко
обрезают, чтобы стимулировать быстрое отрастание молодых вегетативных побегов,
из которых и нарезают черенки; эти побеги характеризуются высокой корнеобразовательной
способностью. Чем сильнее обрезано маточное растение, тем интенсивнее отрастают
новые побеги. В целом успех черенкования в значительной мере определяется
способностью стебля к образованию корней; если же она отсутствует или выражена
слабо, стебель использовать нельзя.
Условия для выращивания черенков
Корнеобразовательная способность свеженарезанных
черенков в сильной степени зависит от температурных условий. Образование корней
определяется химическими процессами; при повышенной температуре скорость
химических реакций возрастает, в результате корни появляются быстрее. Но если в
тепле находится весь черенок, в рост трогается и его верхушка, и какая-то доля
необходимых для роста корня питательных веществ будет поступать уже в верхнюю
часть черенка.
Откуда нарезать черенки
|
Нижний срез узлового черенка должен находиться |
|

Техника черенкования
Листопочковые черенки. Черенки, состоящие из почки с листом, можно нарезать из
любых побегов — зеленых, полу-одревесневших и одревесневших, как вечнозеленых,
так и листопадных растений. Каждый черенок состоит из листа, находящейся в его
пазухе почки и очень короткого отрезка стебля. Из листа поступают вещества,
необходимые для поддержания жизнедеятельности черенка и регенерационных
процессов; из почки вырастает стебель нового растения; первые корни появляются
на небольшом отрезке стебля.

1.
Маточное растение обрезают, чтобы вызвать формирование побегов с высокой
корнеобразовательной способностью.
2.
Отбирают молодой побег с неповрежденным листом и жизнеспособной почкой в его пазухе.
3.
Над самой почкой делают косой срез.

ЛИСТОПОЧКОВЫЙ ЧЕРЕНОК С СУПРОТИВНЫМИ ЛИСТЬЯМИ
1.
Стебель с супротивными листьями срезают над самым узлом и в 2,5—4 см ниже.
2.
Стебелек расщепляют посредине острым ножом. В результате получаются два черенка.
3.
Либо один
лист удаляют. Черенок обрабатывают регулятором роста и
высаживают. Ставят этикетку.

4.
Нижний поперечный срез делают в 2,5—4 см
от верхнего.
5.
Крупный лист подрезают или сворачивают.
Черенок обрабатывают стимулирующим корнеобразование регулятором роста.
6.
Черенок высаживают в горшок так, чтобы
почка была на уровне почвы. Субстрат уплотняют и поливают водой с растворенным
фунгицидом.
Черенки с «пяткой»
Один из традиционных способов размножения —
использование черенков с ≪пяткой≫. Нарезать такие черенки можно из зеленых растущих,
полуодревесневших и одревесневших побегов как вечнозеленых, так и листопадных
растений. Молодой боковом побег отделяют от стебля маточного растения таким
образом, что на его конце остается ≪пятка≫ — кусочек более старой коры и древесины. Смысл подобного
приема заготовки черенков заключается в том, чтобы обеспечить нормальный
дальнейший рост и образование корней в нижней части стебля, а также
предохранить ее от весьма вероятного загнивания. Кроме того, такое утолщенное
основание черенка обладает высокой корнеобразовательной способностью.

1.
Боковой побег берут у основания указательным и
большим пальцами. Резко дергают вниз.
2.
Кончик ≪пятки≫ подрезают, нижние листья удаляют. Основание черенка
обрабатывают регулятором роста.
3.
В земле делают колышком лунку. Черенок
высаживают и поливают водой с растворенным в ней фунгицидом. Ставят этикетку.
Молоткообразные черенки
У черенков молоткообразной формы оставляют
небольшой отрезок стебля более старой древесины, что защищает их от
повреждения гнилями. Этот способ в основном применяется для заготовки
полуодревесневших и одревесневших черенков. Особенно успешно такому
черенкованию поддаются в осенний период многие барбарисы. Для
черенкования лучше всего использовать стебли со слабым ростом, на
которых образуются небольшие боковые побеги.

1.
На маточном стебле над боковым побегом
делают горизонтальный срез.
2.
Нижний срез делают примерно в 2 см от первого.
Листья у основания черенка удаляют.
3.
Нижний срез обрабатывают регулятором роста.
Черенок высаживают, поливают водой с растворенным в ней фунгицидом.
Размножение прививками
|
Прививкой называют соединение двух частей |
|


Прививка глазком. Способ прививки, при
котором в качестве привоя используют почку с кусочком коры и древесины
(глазок), получил название окулировки (от лат. окулус – «глаз», иначе –
прививка глазком). На подвое острым ножом делают Т-образный надрез коры. Края
коры подвоя осторожно отгибают и вставляют глазок. Почка привоя выступает
наружу. Место соединения привоя и подвоя обвязывают. Чаще окулировку производят
в конце лета, но можно это делать и весной. Глазки берут с однолетних побегов.
Выбирают наиболее крупные почки с плодоносящих растений того сорта, который
хотят размножить. В случае удачной прививки, когда обеспечивается срастание
привоя и подвоя, глазок дает начало побегу. Побеги, развивающиеся из почек
подвоя, срезают. Новое растение представляет организм, у которого корневая
система унаследована от подвоя, а почти вся надземная часть — система побегов
привоя.

|
Копулировкой называют сращение черенка с подвоем, имеющим |
|

|
Прививка сближением (аблактировка). Этот способ применяют для трудносрастающихся |
|



Вопросы
для закрепления
1.Перечислите
типы размножения у растений
2.
Что такое способы и каково значение вегетативного размножения растений?
3.В чем сущность и
значение чередования поколений в цикле развития растений?
Контрольные
вопросы по
Разделу 1. «Морфология растений»
1.
Какова роль растений в биосфере и жизни
человека?
2.
Какие органы у растений вегетативные и
какие репродуктивные?
3.
Что такое метаморфоз органов?
4.
Какие функции выполняет корень ?
5.
Какие типы корневых систем вы знаете?
6.
Что такое микориза и какое значение она
имеет для растений?
7.
В чем особенность строения корней бобовых
растений?
8.
Какие части имеет побег?
9.
Что такое почка?
10. Какое
значение имеют покоящиеся, спящие и придаточные почки?
11. Каковы
функции стебля?
12. Каковы
функции и особенности морфологического строения листа?
13. Какие
метаморфозы стебля, листа и побега вы знаете?
14. Какие
жизненные формы бывают у растений?
15. Какие
типы размножения у растений вы знаете?
16. В
чем их сущность?
17. Каково значение вегетативного
размножения растений?
18.В чем сущность и значение чередования
поколений в цикле развития растений?
19.Что такое спорофит и гаметофит?
20.Каково значение цветения в жизни
растений?
21.Как устроен цветок и каковы функции
частей цветка?
22.Что такое опыление?
23.Какие виды опыления существуют в
природе и как приспособлены растения к ним?
24.Как происходит перекрестное опыление?
25.Что такое семя?
26.Каково его строение?
27.Какие условия необходимы для
прорастания семян?
28.Каково происхождение плодов?
29.Для чего они предназначены и какие типы
апокарпных и ценокарпных плодов вы знаете?
30.Какие приспособления обеспечивают распространение семян и плодов у
растений?
6.1.1.
Темы практических работ.
№1.
«Изучение вегетативных органов растения
№2.
Изучение генеративных органов растения.
6.1.2.
Самостоятельная работа
1.Изучение
почек на побегах растений и типов ветвления растений дендросада.
2.Определение
форм листовой пластинки у растений дендросада.
3.Изучение
типов соцветий разных растений Хреновского бора.
4.Провести
морфологический анализ семян овса, фасоли, пшеницы, подсолнечника.
ники Г. Клебс (1857–1918) и К. Гебель (1855–1932). Они фактически сформулировали задачу экспериментальной морфологии, так как считали, что основная задача этого раздела морфологии – научиться управлять индивидуальным развитием растений путем изменения условий существования.
Существенный вклад в экспериментальную морфологию внес Н. П. Кренке (1892–1939), известный своими работами по регенерации и трансплантации у растений и как автор теории циклического старения и омоложения растений. Эта теория заложила научные основы вегетативного размножения растений. Знание закономерностей развития организма, его возрастных изменений позволило Н. П. Кренке уже по ранним стадиям развития давать прогнозы скороспелости растений, что имело большое значение для практики.
Своеобразным продолжением работ морфологов-эволюционистов явились исследования советского физиолога и эколога растений Б. А. Келлера (1874–1945). Основной путь эволюции растений Б. А. Келлер видел в морфолого-физиологической перестройке под влиянием изменяющихся условий среды. Он предложил так называемый метод экологических рядов. Этот метод предполагал изучение постепенной смены условий существования и происходящих при этом постепенных изменений растений.
Итак, приблизительно с середины XIX в. в морфологии, довольно тесно переплетаясь друг с другом, развиваются три направления – срав- нительно-онтогенетическое, сравнительно-филогенетическое (эволюционное) и экспериментально-экологическое. Эти направления исследований почти в равной степени представлены и в настоящее время.
Несмотря на то, что морфология растений наука достаточно древняя, она по сей день не утратила своего значения. Сегодня, как и много веков назад, человечество не перестает волновать проблема удовлетворения своих насущных потребностей и в первую очередь – проблема питания.
Сегодня, как в былые, далекие времена, человек не избавлен от многих тяжелых и коварных недугов, поиски средств лечения которых заставляют его вновь и вновь обращаться к миру растений.
И, наконец, сегодня перед человеком остро и насущно встала проблема, о которой заговорили совсем недавно, но заговорили настойчиво и тревожно – проблема охраны растительного мира, проблема рационального использования его богатств. Эти и многие другие вопросы при-
12
знана решать наука ботаники и ее многочисленные подразделения, в том числе и морфология растений.
Морфология растений как наука имеет научный и прикладной характер. Как научная дисциплина она играет большую роль для систематики и филогении растений, так как только на основании признаков растений можно их отнести к тому или иному таксону и установить родственные связи таксонов различных рангов. Большое значение имеет морфология для генетики и селекции. При проведении генетикоселекционных работ важно иметь сведения по жизнеспособности пыльцы, рыльца пестика, типам опыления и другим показателям, обеспечивающим нормальное осуществление процесса оплодотворения.
Данные морфологии находят широкое практическое применение. Надежным методом в геологии является метод спорово-пыльцевого анализа, основанный на изучении ископаемых пыльцевых зерен и спор, что позволяет определить возраст осадочных пород и целенаправленно вести поиск полезных ископаемых. Этот же метод используется в археологии, геоморфологии, палеогеографиии. Он дает возможность судить о флоре, растительности определенных регионов в далекие геологические эпохи. Спорово-пыльцевой метод находит также применение и в медицине (при выявлении аллергенов), и в товароведении (например, при определении качества меда), и в других областях. Данные морфологических исследований используются в различных отраслях народного хозяйства. Так, подбор пар растений для смешанных посевов (вико-овсяная смесь и др.) основан на изучении урожайности растений в чистых и смешанных посевах. Изучение взаимоотношений некоторых лесообразующих пород с грибами-микоризообразователями способствовало успешному созданию в степной зоне искусственных полезащитных лесных полос, что имело большое значение для развития лесоведения. Не обходятся без морфологических данных и различные ресурсоведческие работы. Именно на основании морфологических исследований процессов размножения можно корректировать сроки и объемы заготовки дикорастущих лекарственных, ягодных, технических и других культур, т. е. давать рекомендации по рациональному использованию естественных природных ресурсов. Только по результатам изучения морфологических особенностей роста и развития растений можно разрабатывать научно обоснованные меры охраны конкретных видов. И, наконец, нельзя забывать о том, что на основании данных морфологии издавна создавались и в настоящее время создаются различные справочные пособия: атласы растений, определители, флоры.
13

НЕКОТОРЫЕ ОСОБЕННОСТИ СТРОЕНИЯ КЛЕТКИ ВЫСШИХ РАСТЕНИЙ
В настоящее время ни у кого не вызывает сомнения, что все живые организмы состоят из клеток. Клетка – это структурный элемент животных и растений.
Изучение клетки начинается с момента ее открытия. В 1665 г. английский естествоиспытатель Роберт Гук (1635–1703), исследуя под микроскопом тонкие срезы пробки, увидел картину, напоминающую пчелиные соты. Эти ячейки, «отверстия или поры», по выражению Р. Гука, он назвал «cellula» – клетка. Наблюдения Р. Гука были продолжены М. Мальпиги и Н. Грю, однако они обращали внимание только на форму клеток, не пытаясь выяснить внутреннее строение. Анатомические исследования растений в XVII–XVIII вв. были немногочисленными и фактически не вносили ничего нового в представление о клетке по сравнению с представлениями Р. Гука, М. Мальпиги и Н. Грю.
Сведения о строении клетки длительное время были отрывочными и очень примитивными. Практически все ботаники XVII – начале XVIII в. представляли клетки как пустоты в растительной ткани, имеющие общие стенки, или как «пузырьки».
Только в 1812 г. немецкий ботаник И. Мольденгауер (1766–1827) применил метод мацерации растительных тканей, получил изолированные клетки и показал, что каждая клетка имеет собственную оболочку. Дальнейшее развитие представление о клетке получило в работе немецкого ботаника Ф. Ю. Ф. Мейена (1804–1830) «Фитотония», вышедшей в 1830 г. Растительную клетку Мейен характеризовал как «пространство, полностью замкнутое и окруженное растительной мембраной».
С 40-х гг. XIX в. начинается изучение внутреннего строения клетки. В 1839 г. чешский ботаник Ян Пуркинье (1787–1869) обнаружил содержимое в животных клетках и назвал его протоплазмой, а в 1846 г. немецкий ботаник Гуго фон Моль (1805–1872) перенес этот термин на содержимое живых растительных клеток. Тем самым Моль подтвердил наблюдения Р. Гука, считавшего, что клетка не пустая, а в ней есть содержимое, которое он называл «соком». Термин протоплазма впоследствии несколько видоизменялся в зависимости от того, какой смысл вкладывали в него исследователи. Так, И. Ганштейн (1822–1880) содержимое одной клетки (без наружной оболочки) называл протопластом. Э. Страс-
14
бургер в 1882 г. предложил термин цитоплазма – обязательная часть клетки, заключенная между плазматической мембраной и ядром.
Постепенно удалось выяснить, что клетка имеет сложное внутреннее строение. Еще в 1676 г. А. Левенгук (1632–1723) фактически обнаружил в клетке пластиды, но назвал их кристаллами. Важное открытие сделал в 1831 г. Р. Броун (1773–1858). При изучении эпидермиса орхидных он дал подробное описание ядра, а в 1842 г. М. Шлейден (1804– 1881) впервые в составе ядра обнаружил ядрышки. В 1882 г. В. Флемминг (1843–1905) в клетках животных обнаружил митохондрии, а в 1904 г. Ф. Мёвес (1878–1923) выявил их в клетках растений. В 1846 г. Моль доказал, что протоплазма растительных клеток обладает способностью к самостоятельному движению. Наблюдения Моля подтвердил в
1850 г. Ф. Кон (1828–1898), а в 1854 г. Н. Принсгейм (1823–1894).
Неоднократно делались попытки изучить образование клеток. Так, в 1835 г. Моль описал деление клеток у водоросли кладофора, а в 1838 г. – деление клеток, из которых впоследствии формировались замыкающие клетки устьиц. В 1841 г. австралийский ботаник Ф. Унгер (1800–1870) наблюдал деление клеток в точке роста растений. Правильное представление о делении клеток имели такие ботаники как Н. И. Железнов, К. Негели (1817–1891), немецкий врач и биолог Р. Реман (1815–1865), немецкий гистолог и эмбриолог А. Келликер (1817–1905), русский зоолог, профессор Московского университета Н. А. Варнек ( 1821–1876). Однако наряду с верными, экспериментально полученными данными об образовании клетки путем деления, существовало ряд неверных представлений об этом процессе. Это касалось и представлений таких признанных в области ботаники и зоологии авторитетов как М. Шлейден и Т. Шванн (1810–1882). Конец всем неправильным суждениям положило меткое высказывание известного немецкого патолога Р. Вирхова (1821–1902): «Omnis cellula e cellula» – «Каждая клетка (происходит только) из клетки».
Таким образом, к 30-м гг. XIX в. уже имелись вполне конкретные представления о строении клетки и в различной форме высказывались предположения, что клетка является основой организации растений и животных. Особенно четко это было сформулировано в клеточной теории, которую Ф. Энгельс наряду с законом превращения энергии и эволюционной теорией Ч. Дарвина назвал одним из трех великих открытий XIX в. Основная заслуга оформления клеточной теории принадлежит Т. Шванну (1839), который использовал собственные данные, а также результаты исследований М. Шлейдена и других ученых.
15
Единство клеточного строения животных и растительных организмов находит подтверждение не только в сходстве строения различных клеток, но, прежде всего, в сходстве их химического состава и метаболических процессов.
Однако, несмотря на наличие общих закономерностей в строении растительных и животных клеток, клетки высших растений имеют ряд специфических особенностей. К таким особенностям относятся наличие прочной клеточной оболочки, наличие пластид и вакуолей.
Клеточная оболочка растительных клеток представляет собой сложное образование, расположенное над плазмалеммой – полупроницаемой мембраной, которая ограничивает цитоплазму клетки и отделяет ее от клеточной оболочки.
Клеточная оболочка – типичный компонент растительной клетки. Благодаря наличию прочной клеточной оболочки сохраняется определенная форма клеток, а протопласт клетки оказывается надежно защищенным.
Оболочка растительной клетки имеет двухкомпонентную структуру. Она состоит из кристаллической основы и матрикса. Скелетную, или кристаллическую, основу клеточной оболочки составляет целлюлоза, или клетчатка. Матрикс состоит из гемицеллюлоз, пектиновых веществ, особого структурного белка (экстенсина) и представляет собой насыщенный водой пластичный гель.
Целлюлоза – основной опорный полисахарид клеточных оболочек растений. В химическом отношении она представляет собой β-1,4-D-глю- кан с эмпирической формулой (C6H10O5)n. Молекулы целлюлозы состоят из остатков D-глюкозы, соединенных β-1,4 гликозидными связями. Количество остатков глюкозы в молекулах целлюлозы варьирует в широких пределах (от 300–500 в вискозных волокнах до 10 000–14 000 в лубяных волокнах), что определяет степень полимеризации ее молекул. Макромолекулы целлюлозы – линейные неразветвленные цепи, которые могут достигать в длину нескольких микрометров, но имеют в толщину всего около 8 нм, что делает их невидимыми даже в электронный микроскоп. Благодаря высокой степени полимеризации молекул целлюлоза не растворяется в воде, органических растворителях, а также в слабых растворах кислот и щелочей.
В оболочке растительной клетки молекулы целлюлозы составляют сложные аггрегации. Приблизительно 40–60 макромолекул целлюлозы, располагаясь параллельными тяжами, образуют мицеллы. Молекулы целлюлозы в мицеллах связаны таким образом, что формируют кристал-
16
лоподобные структуры. Благодаря этому для мицелл (часто говорят, для клеточной оболочки) характерно свойство анизотропии, что обеспечивает двойное лучепреломление. Существование мицелл установлено с помощью рентгеноструктурного анализа.
Мицеллы образуют микрофибриллы, или мицеллярные пучки. В состав каждой микрофибриллы входит около 160 мицелл. Толщина микрофибрилл 35–100 нм, а длина достигает 500–600 нм. Несколько сотен микрофибрилл составляют фибриллы. Фибриллы – довольно толстые образования диаметром от 300 до 1500 нм, что позволяет их видеть с помощью светового микроскопа.
Фибриллы погружены в матрикс. Основу матрикса клеточной оболочки составляют гемицеллюлозы и пектиновые вещества. Гемицеллюлозы – группа полисахаридов, молекулы которых образованы остатками различных сахаров. Чаще всего они представлены остатками молекул гексозных сахаров (глюкоза, галактоза, моноза, фруктоза) и имеют эмпирическую формулу аналогичную формуле глюкозы – (C6 H10O5)n. Однако в отличие от молекул целлюлозы молекулы таких гемицеллюлоз (гексозанов) менее полимеризированы, они включают не более 200 (от 50 до 200) моносахаридных остатков. Гемицеллюлозы могут быть образованы также остатками пентозных (рибоза, арабиноза) сахаров (пентозаны) и тогда их формула (C5H8O4)n. Молекулы гемицеллюлоз могут быть линейными или разветвленными. Особенности строения гемицеллюлоз отражается на их химических и физических особенностях. В отличие от целлюлозы, которая не способна впитывать воду, гемицеллюлозы способны набухать в воде, но, как и целлюлоза, не растворяются в ней и органических растворителях. В то же время гемицеллюлозы растворяются в слабых растворах щелочей и легко гидролизуются в кислотах.
Пектиновые вещества – кислые высокомолекулярные полисахариды растений. По химической природе пектиновые вещества представляют собой неразветвленные полимеры галактуроновой кислоты, в которой часть карбоксильных групп этерифицирована метиловым спиртом. В состав пектиновых веществ входят также нейтральные моносахариды – галактоза, рамноза, арабиноза, ксилоза. Пектиновые вещества, входящие в состав матрикса клеточной оболочки, способны сильно набухать в воде и образовывать гель. Некоторые из пектиновых веществ могут растворяться в воде, кроме того, они легко разрушаются под действием кислот и щелочей.
Полисахариды матрикса не просто заполняют промежутки между мицеллами целлюлозы. Они располагаются в оболочке клетки довольно
17
упорядоченно и образуют многочисленные ковалентные или водородные связи как друг с другом, так и с микрофибриллами.
Образование клеточной оболочки связано с делением клетки (цитокинезом). В конце телофазы митоза в экваториальной зоне делящейся клетки между нитями ахроматинового веретена (веретена деления, митотического веретена) образуется фрагмопласт. Впервые эту структуру обнаружил и описал немецкий ботаник Л. Еррера в 1888 г. и указал, что она имеет бочонковидную форму и состоит из фибрилл. В настоящее время установлено, что фрагмопласт представляет собой систему микротрубочек. Микротрубочки – это лишенные мембраны органеллы, представляющие собой цилиндрические образования диаметром около 24 нм, длина которых может достигать нескольких микрометров. В центре микротрубочка имеет полость, а ее стенка образована 13 рядами димерных комплексов глобулярных белков – тубулинов. Микротрубочки располагаются преимущественно в периферических слоях цитоплазмы. При делении клетки веретено деления и фрагмопласт оказываются тесно связанными друг с другом благодаря наличию в них общих микротрубочек. Фрагмопласт обнаружен у всех высших растений и зеленых водорослей, за исключением представителей класса Chlorophyceae.
Разделение материнской клетки на две дочерние начинается с формирования клеточной пластинки, в образовании которой принимают участие диктиосомы. Совокупность всех диктиосом составляет комплекс Гольджи (аппарат Гольджи). Диктиосомы – это одномембранные органеллы, состоящие из плоских округлых цистерн. В состав диктиосом растений входит от двух до семи (реже больше) цистерн, расположенных одна над другой в виде стопки. Между отдельными цистернами стопки имеются небольшие промежутки, заполненные фибриллярными и трубчатыми элементами, функция которых неизвестна. Диктиосомы выполняют секреторную функцию. В них происходит энергичное образование пузырьков (пузырьки Гольджи, тельца Гольджи), в которых содержатся секретируемые вещества – полисахариды или полисахаридно-белковые комплексы, обладающие высокой вязкостью. Вязкость содержимого пузырьков Гольджи объясняется наличием большого количества пектиновых веществ, за что их часто называют пектиновыми пузырьками.
Пектиновые пузырьки мигрируют в область фрагмопласта, выстраиваются в горизонтальном направлении в его центральной части, сливаются и начинают формировать клеточную пластинку. На первых стадиях развития клеточная пластинка закладывается в виде диска в центральной части фрагмопласта. Пектиновые пузырьки неплотно прилега-
18
ют друг к другу. При слиянии пузырьков в клеточной пластинке остаются небольшие промежутки – плазмодесменные каналы, выстланные плазмалеммой. В них проходят плазмодесмы – мельчайшие цитоплазматические тяжи, соединяющие протопласты дочерних клеток. Постепенно, по мере того как в центральной части фрагмопласта микротрубочки исчезают, на его периферических концах появляются новые и фрагмопласт расширяется, направляясь к стенкам делящейся материнской клетки. Вместе с ним расширяется и клеточная пластинка, формирование которой идет от центра к периферии. Когда клеточная пластинка достигнет оболочки делящейся клетки, процесс деления фактически завершается. Образовавшиеся дочерние клетки должны достичь определенных размеров (вырасти) и сформировать собственные оболочки.
В образовании клеточной оболочки принимают участие комплекс Гольджи и плазмалемма клетки. В период роста клетки формируется первичная клеточная оболочка. На первых этапах развития оболочка дочерних клеток состоит из остатков оболочки материнской клетки и участков вновь образующейся первичной клеточной оболочки. Первичная клеточная оболочка начинает формироваться между клеточной пластинкой и плазмалеммой. Как полагают, гемицеллюлозы и пектиновые вещества доставляются к месту синтеза оболочки пузырьками Гольджи, а образование целлюлозы связано с деятельностью плазмалеммы. Первичная клеточная оболочка довольно пластичная. В ее состав входит около 15% целлюлозы, остальная часть приходится на долю матричных компонентов. Встраивание молекул целлюлозы в клеточную оболочку может происходить двумя способами: путем интуссусцепции и путем аппозиции. При интуссусцепции новые молекулы целлюлозы внедряются между уже встроившимися в оболочку молекулами. При оппозиции новые молекулы целлюлозы накладываются изнутри на уже отложенные элементы клеточной оболочки. В первичной клеточной оболочке, как и в клеточной пластинке, есть участки, через которые проходят плазмодесмы. Эти участки первичной клеточной оболочки, пронизанные плазмодесмами и более тонкие, чем остальные, называются первичными поровыми полями.
Так как в первичной клеточной оболочке преобладают гемицеллюлозы и пектиновые вещества, содержится много воды, а молекулы целлюлозы расположены неупорядоченно, оболочка способна растягиваться, а клетка может расти как в длину, так и в ширину.
Характер роста определяет будущие морфологические особенности сформированной клетки. Если клетка более или менее равномерно раз-
19
растается в длину и ширину, она становится изодиаметричной (с равными продольным и поперечным диаметром) или таблитчатой (один из диаметров не более чем в два раза, превышает другой). Такие клетки называются паренхимными. Если длина значительно превышает ширину, клетки называются прозенхимными.
Первичные клеточные оболочки характерны для клеток меристемы, для молодых растущих клеток, а также для клеток некоторых постоянных тканей.
У большинства клеток, достигших своих предельных размеров и прекративших рост, над первичной клеточной оболочкой путем аппозиции начинает формироваться вторичная клеточная оболочка. В состав вторичной клеточной оболочки входят те же компоненты, что и в состав первичной, но соотношение полисахаридов в ней иное. Большая часть (до 60 %) приходится на долю целлюлозы, а во вторичных оболочках лубяных волокон льна содержание целлюлозы может достигать 95 %. Кроме того, увеличивается степень полимеризации ее молекул, упорядочивается расположение микрофибрилл в матриксе. Объем матрикса и количество воды значительно уменьшается, но связи между компонентами вторичной клеточной оболочки усиливаются.
Для вторичной оболочки характерна слоистость. Она возникает в процессе ее формирования и связана с расположением в ней микрофибрилл целлюлозы. Наиболее часто вторичная оболочка состоит из трех слоев, которые различаются ориентацией микрофибрилл. В каждом слое микрофибриллы располагаются строго параллельно друг другу, но угол наклона микрофибрилл в каждом слое по отношению к предыдущему меняется. Вторичная клеточная оболочка не сплошная. В тех местах, где в первичной оболочке были сформированы первичные поровые поля, вторичная оболочка прерывается. Перерыв во вторичной клеточной оболочке, расположенный над первичным поровым полем, называется порой. Поры в соседних клетках образуются одна против другой, т. е. возникает пара пор. Эти поры соединяются поровыми каналами. Поры бывают простыми и окаймленными.
В процессе жизнедеятельности растения вторичная клеточная оболочка может претерпевать вторичные изменения: одревеснение, опробковение, кутинизацию, минерализацию, ослизнение.
Одревеснение, или лигнификация (от лат. lignum – древесина), – процесс, связанный с отложением в матриксе вторичной клеточной оболочки лигнина – сложного полимерного соединения, химическое строение которого окончательно не установлено. Лигнин – один из важней-
20
ших компонентов вторичных клеточных оболочек – образуется только в оболочках клеток высших растений (за исключением мохообразных). Он откладывается в матриксе оболочки некоторых специализированных клеток (склеренхимы, трахеид, члеников сосудов) между микрофибриллами и значительно повышает ее прочность. В то же время лигнин делает оболочку не способной пропускать воду с растворенными в ней минеральными веществами, нарушает газообмен и, в конечном счете, способствует отмиранию живого содержимого клетки. Одревесневшие клеточные оболочки обладают ультраструктурой, которую можно сравнить со структурой железобетона.
Из жироподобных веществ во вторичных оболочках клеток некоторых тканей (феллемы, эндодермы) откладывается суберин (от лат. suber – пробка). Суберин не растворяется в органических и неорганических растворителях, в кислотах, но разрушается в концентрированных щелочах. Суберин может откладываться в клеточной оболочке в виде отдельных пластинок или в виде сплошного слоя. В последнем случае протопласт оказывается в полной изоляции от внешней и внутренней среды и отмирает.
По химической природе к суберину близки гидрофобные полимеры кутин и воск. Кутин откладывается на поверхности клеточной оболочки, образуя тонкую пленку – кутикулу. Кутикула выполняет защитную функцию и предохраняет надземные органы растений от излишнего испарения воды.
Аналогичные функции выполняет и восковой налет, развивающийся на поверхности различных органов растения. В отличие от кутина и суберина воск растворяется в органических растворителях и легко плавится. Воск откладывается в виде аморфного или кристаллического слоя и выполняет защитную функцию. Воск может также входить в состав кутикулы, что усиливает ее барьерные свойства.
Минерализация клеточных оболочек обусловлена отложением в них кремнезема или солей кальция (оксалата, карбоната). Кремнезем в аморфном состоянии откладывается в оболочках эпидермальных клеток некоторых растений (злаков, хвощей, волосков крапивы). Оксалат и карбонат кальция могут откладываться в клеточных оболочках или образуют в клетках цистолиты и различные кристаллы.
Ослизнению оболочек способствует наличие в них большого количества гемицеллюлоз и особенно пектиновых веществ. Ослизнение сопровождается связыванием большого количества воды, что приводит к образованию на поверхности слизей, представляющих собой смесь поли-
21
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
БИОЛОГИЯ
УДК 575.85.827 А.Г. Юсуфов
История развития экспериментальной морфологии растений (методы, методология и последствия)
Дагестанский государственный университет, plantphysiologist@,mail.ru
Европейские ботаники в XIX и XX вв. пытались объяснить природу действия комплекса внешних факторов на морфологические особенности растений. Эта область была выделена как экспериментальная морфология растений. Изменчивость морфологических признаков растений при этом трактовалась в рамках идей Ламарка.
Развитие генетики опровергло эти идеи. Результаты исследований ранних ботаников говорят лишь о роли нормы реагирования генотипа на воздействия. Выяснение механизмов действия факторов внешней среды должно соответствовать генетическим подходам.
Ключевые слова: растения, экспериментальная морфология растений, эволюция, норма реакции генотипа, модификации.
European botanists in 19 and 20-th centuries made attempts to explain the nature of effects of environmental factors. Thus was formed a field of research called «experimental plant morphology». Variability of morphological features of plants was treated in the framework of the ideas of Lamarck.
Development of genetics disproved these ideas. Early botanical works tell only on the norm of reaction of genotype to changes of environmental conditions. To elucidate the mechanisms of actions of environmental features it is need to use genetics methods.
Keywords: plant, experimental morphology, Lamarck, evolution, norm reaction of genotype, modifications.
Морфология животных и растений — один из древнейших разделов науки с широким кругом вопросов. Данные морфологии используются в систематике, филогенетике, экологии, генетике и для познания функциональных особенностей организма. Морфология — основа биологии, изучает форму (габитус) живых существ и отдельных их органов с точки зрения реализации генетической программы, что проявляется в специфике организации индивидуумов разных видов и их органов.
Экспериментальная же морфология занималась изучением факторов, способствующих изменению габитуса, сроков цветения и продолжительности жизни и индивидуальности растений. Это направление еще называют биологией развития растений. Область ботаники, сложившуюся на стыке морфологии и физиологии растений в конце XIX и начале XX вв., занимающуюся «разоблачением тайны образования растительных форм», К.А. Тимирязев предложил выделить как новый раздел биологии — экспериментальную морфологию растений. Он восторженно писал, что это направление, «пробиваясь отдельными струйками в XIX в., сольется в широкий поток уже за порогом двадцатого». Позднее в работах «Ч. Дарвин и полувековые итоги дарвинизма» и «Столетние итоги физиологии растений» [3] он констатировал, что его ожидания полностью оправдались на примере исследований начала ХХ в. [4; 8; 9]. Однако до сих пор результаты и последствия начального периода развития экспериментальной морфологии растений, о чем с восхищением писал К.А. Тимирязев, остаются без анализа. Этому вопросу и посвящено сообщение.
В качестве факторов, непосредственно влияющих на изменение формы растений, К.А. Тимирязев указывал свет, температуру, влажность, земное притяжение, опираясь на данные Лева-ковского, Аскенази, Визнера, Фехтинга, Бонье и др. Основы экспериментальной морфологии, однако, были заложены еще ранее при изучении связи роста растений с земным притяжением (Найт) и условиями освещения (Декандоль). Исследования указанных авторов были дополнены позднее наблюдениями о приуроченности определенных форм растений к соответствующим географическим областям Земли (Варминг).
Уместно поставить вопросы: почему К. А. Тимирязев был так увлечен достижениями экспериментальной морфологии растений? каково последующее их влияние на развитие ботаники? В конце XIX начале ХХ вв. было характерно критическое отношение к учению Ч. Дарвина, и этот период обозначен как период его «отрицания» [5]. Данные генетики (учения о мутационной изменчивости и чистых линиях) поставили под сомнение роль отбора в эволюции. Фундаментальные положения дарвинизма пытались опровергнуть такими данными, «против естественного отбора была проведена атака» со всех сторон [2, с. 216].
Зарождающаяся генетика со своими результатами изучения изменчивости и наследственности начала подрывать устои учения об отборе. Для их защиты К.А. Тимирязев [2] в статье «Ч. Дарвин и полувековые итоги дарвинизма» использовал данные исследований Уэльдона, Бербанка и экспериментальных морфологов растений. Экспериментальные данные начального периода становления генетики были посвящены изучению причин изменчивости, о которых Ч. Дарвин мало знал. Попытки же К.А. Тимирязева подкрепить учение Дарвина ссылками на результаты экспериментальной морфологии растений не оправдались в связи с защитой представлений о наследовании благоприятных изменений, т. е. идей Ж.Б. Ламарка и механоламаркистов.
Ж. Ламарк акцентировал внимание на зависимости растительных форм от влияния среды, и по этой причине его «учение более привилось в ботанике, чем в зоологии» [3]. По словам К.А. Тимирязева, ламаркизм, тем не менее, неспособен ответить на основной вопрос, почему эти формы так совершенны, так целесообразны? Он нередко переоценивал достижения экспериментальной морфологии растений для развития дарвинизма. Так, касаясь вопроса об источниках изменчивости, указывал, что «орудием (источником. — А.Ю.) изменчивости служит действие среды», о чем якобы энергично Ч. Дарвин высказался также в письме к Гексли: «Кой черт изменяет формы, если это не внешние условия?»
Все последующее развитие науки, продолжает К.А. Тимирязев, было подтверждением верности этого воззрения. Несколькими страницами раньше читаем: «… возражение, что дарвинизм … упирается в тайну явления изменчивости» разбивается фактами, полученными в области физиологии растений [1, с. 517]. Далее: «… из того факта, что приспособления вырабатываются как бы автоматически под влиянием окружающих факторов, еще не вытекает обратное заключение, что действие этих факторов само по себе всегда целесообразно …» [1, с. 518]. Эти выдержки характеризуют противоречивость оценок А.К. Тимирязевым достижений в области экспериментальной морфологии растений для развития дарвинизма.
Чтобы разобраться в этом вопросе, обратимся к анализу некоторых типичных данных того периода. Еще в работах Ламарка было выдвинуто положение о возможности получения наследственных уклонений путем выращивания растений в определенных условиях. Ж.Б. Ламарк вообще не признавал изменений, не передающихся по наследству, впервые поставил эволюционный процесс в зависимость от наследования благоприятных изменений, что впоследствии еще более рельефно выступало в работах Э. Геккеля и Г. Спенсера [6]. Это выразилось в известном утверждении Г. Спенсера: «Либо была наследуемость приобретенных свойств, либо вовсе не было эволюции (развития)». Подобные представления оказались настолько соблазнительными, что даже А. Вейсман в своих ранних работах не удержался от признания возможности наследования величины семян, измененной в поколениях под влиянием климата [6, с. 93]. Э. Радль [7] подчеркивал, что «доказательством» идеи о возможности наследования приобретенных под влиянием условий среды изменений особенно увлекались французские ботаники конца прошлого и начала нынешнего веков. При этом они опирались на наблюдения по переносу растений с гор на равнину или с равнины на горы. В таких опытах были отмечены морфологические (высота и габитус растений, форма и жилкование листа, величина семян) и биологические (сроки цветения, продолжительность жизни) изменения, которые сохранялись в поколениях при наличии индуцирующих условий. При возвращении растения в прежние и типичные для них условия восстанавливалась характерная их норма.
Обращалось внимание и на оценку изменчивости явлений морфогенеза при реализации процессов регенерации и нарушении полярности (ориентации) у растений, которые якобы проливали свет на обсуждаемые вопросы [1, 2]. Исследования конца XIX в. по изучению влияния разных физических факторов на развитие и продолжительность жизни растений, во-первых,
показали возможность значительного смещения таким путем указанных свойств [8, 9], во-вторых, послужили основанием для недооценки роли наследственности в явлениях развития и морфогенеза [10]. Это второе направление в конечном итоге было связано с защитой идеи о наследовании благоприобретенных свойств и врожденной способности растений к прямым приспособлениям и «самоприспособлениям» — основные постулаты механоламаркистов-ботаников (Бонье, Константен, Веттштейн, Варминг и др.). Механоламаркизму в целом было присуще признание возможности возникновения целесообразной организации «путем прямого или функционального приспособления», как результата суммирования ответных реакций организма на внешние воздействия [5, с. 173]. Поэтому эти данные без основания использовались «для подкрепления механоламаркистской доктрины эволюции» [5, с. 195-197]. Современная биология объединяет факты географической и онтогенетической изменчивости растений, исходя из представлений о норме реагирования генотипа. Норма реагирования растений (представления о пределах изменчивости индивидуумов в зависимости от условий культивирования) составляет предмет изучения генетики и теории эволюции. Наследуются в поколениях не признаки, а норма реакции генотипа. Поэтому изменения морфологических признаков растений ограничены возможными пределами реагирования их генотипов на условия культивирования, что проявляется в виде модификаций в облике фенотипов. Существенные изменения нормы реагирования на условия происходят только после преобразованний программы генотипа. Изменения показателей морфогенеза в зависимости от условий культивирования, силы земного притяжения и реализации процессов регенерации различны и свидетельствуют о возможных пределах вариации нормы реакции генотипов разных растений.
В указанный период было отмечено усиление исследований в области регенерации животных и растений, чему отчасти способствовали идеи Г. Дриша [12]. Удалось выяснить структурные изменения и условия, благоприятствующие восстановлению целостности изолированных частей и организма у животных и растений проявлением способности их регенерации. Эти исследования привели и к постановке некоторых общих вопросов. В дискуссиях по вопросу о происхождении и эволюции явлений регенерации, которые проходили в конце XIX в. [11], ботаники ограничились признанием их целесообразности. Такая постановка вопроса была общей и отвечала духу идей Г. Дриша [12] об универсальном их значении для живых существ. Ссылки на признание роли естественного отбора в реализации процессов регенерации [4] не могли изменить уже сложившуюся общую ситуацию.
К.А. Тимирязев касался данных о регенерации при оценке значения внешних условий для регуляции морфогенеза и развития растений, а также при рассмотрении явлений наследственности. Говоря о размножении бегонии листьями, он писал: «стоило ботанику (Сидней-Вайнзу) … произнести одно слово — бегония, чтобы разрушить учение о двух плазмах — зародышевой и телесной [1, с. 522]. Теперь очевидно другое, что такие исследования принесли только подтверждение хромосомной теории наследственности [13]. Именно они отчасти объяснили вопрос, волновавший К.А. Тимирязева, каким образом «смертная плазма родит бессмертную носительницу наследственности», исходя из закономерностей расхождения хромосом при делении соматических клеток.
Вопрос о происхождении и эволюции явлений регенерации растений до сих пор не решен. Решение его связано с необходимостью изучения восстановления целостности организма и отдельных его структур путем реализации механизмов молекулярной репарации и деления клеток, выяснения условий, благоприятствующих их осуществлению. Эволюция процессов регенерации у растений происходила путем соотбора их с особенностями, определяющими экологическое приспособление, что привело к возникновению неоднозначных способов восстановления целостности. Причины дивергенции видов по способности к регенерации могут быть разнообразными, и не всегда можно найти их эволюционную интерпретацию [15].
На исследования по регенерации растений оказали влияние ламаркистские идеи, характерные для экспериментальной морфологии. Уже в ранних работах [4] ставился вопрос о возможности усиления способности к регенерации при повторных повреждениях (Дарвин) и размножениях растений черенками. Но этот вопрос не был решен и позднее, хотя иногда допускалась возможность «прямого приспособления» семенного потомства из черенков передачей свойства
повышенной активности к регенерации таким путем размноженных в ряде поколений растений [14]. Все это было результатом методологически непродуманных опытов.
При последовательных посадках для укоренения исходных черенков одно- и многолетних растений после очередного появления корней происходит измельчание вегетативного потомства благодаря задержке роста из-за энергетического истощения тканей в результате повторных посадок [15]. Тем не менее, семенное потомство однолетних растений по вариантам 2 и 3 «поколений» черенков проявляет высокую жизнеспособность и укореняемость не из-за передачи ему способности к укоренению исходного вегетативного потомства, а благодаря его омоложению. У многолетних растений (бересклет японский и самшит обыкновенный) 2 и 3 «поколения» черенков на 2-й год по приросту не уступают потомству черенков первой посадки [15].
В связи с изучением потомства вегетативного происхождения остро встал вопрос о последствиях регулярных способов такого размножения. Ведет ли постоянное вегетативное размножение к вырождению клона и вида в ряду поколений? Для сторонников ламаркизма такой исход не вызывал сомнений с учетом наблюдений ускоренного наступления плодоношения у плодовых в случае прививки черенков зрелых индивидуумов и малой устойчивости к неблагоприятным факторам клоновых насаждений вегетативно размножаемых растений. Эти явления были использованы для доказательства передачи семенному потомству онтогенетического состояния развития вегетативного поколения маточных растений [14; 16].
Известно, что Ч. Дарвин высоко оценивал роль полового размножения в эволюции, что подтвердилось последующим развитием биологии из-за наличия механизмов повышения изменчивости в поколениях. В то же время очевидна и эволюционная устойчивость растений, воспроизводящихся в природе вегетативно без признаков вырождения [15]. Отбор поддерживает разные формы размножения, повышающие надежность онтогенеза растений и конкурентную способность вида. Многие вегетативно возобновляемые сорняки проявляют устойчивость даже при жесткой агротехнике, доставляя немалые хлопоты полеводам. К тому же, для некоторых видов вегетативное размножение в природе стало доминирующим способом закрепления уже занятой территории. При этом происходят не качественные изменения генетической информации, а количественный рост ее части, уже апробированной по устойчивости в данных условиях. В критических условиях отмирание общей массы растений с одинаковой наследственностью у насаждений вегетативного происхождения наступает не от старческого их вырождения в поколениях, а в результате наследственной неустойчивости такого потомства к изменившимся факторам.
Известно также, что вегетативное размножение играет большую роль в жизни многих видов. Оно усиливает их устойчивость путем закрепления гетерогенности и полиплоидии, удержания занятой территории плотным ее насыщением и воспроизведением потомства в условиях ограничения полового процесса [15].
Эксперименты в области регенерации растений, проведенные в конце XIX — начале ХХ вв., имели значение для понимания роли функционального состояния в изменении структуры и продолжительности жизни. Серией убедительных опытов на изолированных листьях разных растений было показано постепенное приобретение черешком после укоренения и по мере увеличения сроков культивирования структурных особенностей, характерных для стебля [17]. Речь шла о возможности постепенного «преобразования» черешков в стеблеподобные образования. Такие онтогенетические данные представители экспериментальной морфологии рассматривали в связи с филогенетическими событиями — наследованием приобретенных изменений.
Изучение вопроса продолжительности жизни растений представляется методически трудной задачей. Поэтому укорененные листья в этом отношении быстро привлекли внимание как удачная модель для анализа влияния условий культивирования на увеличение продолжительности их жизни. Многие экспериментальные морфологи при изучении продолжительности жизни все сводили к влиянию внешних условий. Поэтому заслуживают одобрения попытки акцентирования внимания на роли наследственности в ее определении у индивидуума [18]. Несмотря на это, поздние отечественные ламаркисты стали на путь принижения роли наследственности в развитии и продолжительности жизни индивидуума. Ч. Дарвин подчеркивал, что долговечность индивидуума у растений разных филогенетических групп эволюционно фиксируется в связи с частотой воспроизведения их потомства. Сроки старения и смерти индивидуу-
мов у растений закреплялись отбором не самостоятельно, а ради воспроизведения нового потомства [15]. Приспособительное значение старения и отмирания индивидуумов обусловлено отбором особей по темпам воспроизведения потомства, что является результатом приспособления растений.
В итоге можно сказать, что исследования в области экспериментальной морфологии в конце XIX — начале ХХ вв., безусловно, имели значение для анализа онтогенетических причин становления формы, они стимулировали развитие экспериментальной ботаники в последующем [19]. Однако на начальных этапах представители экспериментальной морфологии растений все же отстаивали положения ламаркизма. Эта позиция еще долго сохранялась и давала о себе знать своими отрицательными последствиями, что хорошо известно в прошлой нашей действительности. Поэтому в целом влияние их для развития ботаники и эволюционной теории было порой даже отрицательным. Ранние и поздние исследования в области экспериментальной морфологии не раскрыли причины изменчивости растений, на что возлагал большие надежды К.А. Тимирязев. К тому же, опираясь только на ее методы, они не могли быть понятыми.
В связи с оценкой истоков развития экспериментальной морфологии растений следует подчеркнуть отсутствие у ее сторонников методических возможностей для проведения таких исследований из-за трудности регулирования конкретных факторов морфогенеза. Наблюдения, касающиеся изменения морфогенеза растений при переносе их с равнины на горы или обратно и с географическим распространением видов, не могли служить основанием для широких обобщений. Они свидетельствовали о роли воздействия комплекса, а не отдельных факторов, в морфогенезе. Возможности регулирования и оценка действия факторов появились лишь позже. Для этого необходимо было утверждение генетического подхода в морфологии, против чего как раз выступили многие ее представители. По этой причине имидж К.А. Тимирязева среди наших генетиков оставляет желать лучшего.
Только с развитием генетического подхода стало проясняться и эволюционное значение данных, полученных ранними экспериментальными морфологами растений. С позиций оценки роли модификаций в онтогенезе и широкой нормы реакции генотипов в приспособлении вида к неблагоприятным условиям можно объяснить биологический смысл изменений в структуре, ритме развития и продолжительности жизни растений в разных условиях.
Методы начального этапа развития экспериментальной морфологии растений не позволяли изучить природу морфогенеза, конкретизировать роль отдельных внешних факторов в реализации программы морфогенеза. Методология же ее оказала отрицательное влияние на развитие исследований в этой области.
Сейчас ведутся дискуссии о создании теоретической биологии, ее задачах и методах. В этой области издается специальный журнал (J. of theoretical biology), публикуются разные сборники и статьи. Однако пока нет общепринятого определения ее предмета и задач. Поэтому нередко ставится под сомнение возможность создания теоретической биологии, соответствующей по значимости теоретической физике. В случае положительного решения этого вопроса одной из важных ее задач останется познание природы, способов и механизмов реализации генетической программы морфогенеза и развития растений и животных [20; 21; 24]. Существуют попытки компьютерного моделирования последовательных этапов морфогенеза растений [22; 25], результаты которых нуждаются еще в экспериментальном подтверждении.
Литература
1. Тимирязев К.А. Исторический метод в биологии // Избр. соч. — Т. 3. — М.: Сельхозгиз, 1949. — С. 352-600.
2. Тимирязев К.А. Ч. Дарвин и полувековые итоги дарвинизма // Избр. соч. — Т. 4. — М.: Сельхозгиз, 1949. — С. 49-234.
3. Тимирязев К.А. Столетние итоги физиологии растений // Избр. соч. — Т. 2. — М.: Сельхозгиз, 1948.- С. 321-406.
4. Goebel К. Einleitung in die experimentell Morphologie der Pflanzen. — Leipzig und Berlin, 1908. — 241 S.
5. Завадский К.М. Развитие эволюционной теории после Дарвина. — Л.: Наука, 1973. — 423 с.
6. БляхерЛ.Я. Проблемы наследования приобретенных признаков. — М.: Наука, 1978. 116 ВЕСТНИК ДАГЕСТАНСКОГО ГОСУДАРСТВЕННОГО УНИВЕРСИТЕТА. 2012. ВЫП. 1
7. Radl E. Geschichte der biologischen Theorien. Bd. II. — Leipzig, 1909. — S. 400-420.
8. Константен Ж. Растение и среда (Приспособление-эволюция). — М., 1908. — 341 с.
9. Клебс Г. Произвольное изменение растительных форм. — М., 1905. — 115 с.
10. Лысенко Т. Д. Агробиология. — М.: Сельхозгиз, 1952. — 778 с.
11. ВоронцоваМ.А. Регенерация органов у животных. — М.: Советская наука, 1949. — 270 с.
12. Дриш Г. Витализм (его история и система). — М.: Наука, 1915. — 260 с.
13. Batteson W. Heredity and variation on Modern Lights // Darvin and Modern Science. — Cambridge, 1910. — P. 85-101.
14. Есаян Г.С. Укореняемость черенков семенного потомства вегетативно размножаемых гибридов томата // ДАН СССР. — 1950. — Т. 71, № 6. — Махачкала, — С. 1120-1124.
15. Юсуфов А.Г. Механизмы регенерации растений. — Ростов н/Д: Изд. РГУ, 1982. — 172 с.
16. Есаян Г.С. Старение растений томата при вегетативном размножении // Журн. общ. биол. — 1950. — Т. 11, № 4. — С. 274-297.
17. Winkler H. Uber die Umwandlung des Blattstiele zum Stengel // Jahrb. wiss. Bot. — 1908. -Bd. 45, H. 1. — S. 1-83.
18. Кренке Н.П. Теория циклического старения и омоложения растений и практическое ее применение. — М.: Сельхозгиз, 1940. — 134 с.
19. Манойленко К.В. Развитие эволюционного направления в физиологии растений. — Л.: Наука, 1974. — 254 с.
20. Poething R.S. Phase and the regulation of shoot morphogenesis in Plants // Sci. — 1990. -V. 250. — Р. 223-230.
21. Белоусов Л.В. Биологический морфогенез. — М.: МГУ, 1987. — 239 с.
22. Торнли Дж. Г.М. Математические модели в физиологии растений. — Киев, 1982. — 310 с.
23. Чуб В.В. Роль позиционной информации в регуляции развития органов цветка и листовых серий побега. — М.: Бином, 2010. — 264 с.
24. Positional control in plant development / Eds. Barlow P.W., Carr, D.J. — Cambridge: Univ. press, 1984. — P. 349-402.
Поступила в редакцию 1 ноября 2011 г.
2.1 Макроскопические изменения морфологии растений .
а) некрозы — отмирание ограниченных участков ткани, довольно специфичны; следует различать :
— точечные и пятнистые некрозы — отмирание тканей в виде точек и пятен; при действии озона,
— межжилковые некрозы — отмирание листовой пластинки между боковыми жилками первого порядка (при воздействии SO2);
— краевые некрозы — характерные, четко различимые формы у лип под действием поваренной соли, применяемой для таяния льда; сочетание межжилковых и краевых некрозов приводит к появлению узора типа “рыбьего скелета”;
— верхушечные некрозы — характерны для хвойных, темно-бурые, резко отграниченные некрозы кончиков хвои под воздействием SO2 или обесцвеченные — под влянием HF.
При развитии некрозов сначала наблюдаются изменения в окраске, затем после гибели клеток пораженные участки оседают, образуются разрывы листовой пластинки.
б) преждевременное увядание — под действием этилена в теплицах, цветки гвоздики при этом не раскрываются, увядают лепестки орхидей;
в) опадение листвы (дефолиация) — в большинстве случаев появляется после появления хлорозов и некрозов; примерами может служит уменьшение продолжительности жизни хвои, ее осыпание у ели, преждевременное опадание листвы у липы, конских каштанов, у крыжовника и смородины под действием SO2; дефолиация приводит к уменьшению ассимилирующей площади, сокращению прироста, растормаживанию почек и преждевременному образованию новых побегов;удобным объектом исследования в таких случаях являются хвойные породы, имеющие многолетнюю хвою. В чистых природных экосистемах средний возраст хвои сосны составляет у южной формы 3-4 года, у северной — 8-9 лет, хвои ели — 7-8 лет. При воздействии токсичных газов возраст хвои сокращается, так что в отдельных случаях на деревьях остается только однолетняя хвоя
Примеры применяемых на практике тест-растений.
Табак Bel W3. Этот сорт выведен специально для биоиндикации озона. Уже при слабом воздействии О3 через несколько дней на листовой пластинке густо образуются некротические пятна серебристого цвета. Этот сорт хорошо зарекомендовал себя в ряде стран и нашел более широкое применение по сравнению с другими индикаторами (виноград, фасоль, шпинат, крапива, петуния).
Метод заключается в экспонировании растений в различных условиях местообитания и определении степени поражения листовых пластинок хлорозом. При подготовке исследований должна быть строго соблюдена методика выращивания табака. Растения выращиваются на гидропонике с питательным раствором при температуре 24°С и рН 5,5-6,0. На каждый участок помещается по 72 экземпляра табака. При этом обеспечиваются достаточный полив и затенение на 40-50%. По оценочным шкалам определяется интенсивность хлороза в баллах, которая затем служит материалом для составления карт полей озона. Методика широко используется в Западной Европе — с помощью этого метода составлены карты загрязнения озоном крупных регионов
Некрозы хвои и продолжительность ее жизни у сосен, поврежденных дымом. Особенно удобными параметрами являются верхушечные некрозы хвои и проложительность ее жизни; их можно определять круглогодично, на основании этих параметров создают картосхемы зон газодымового стресса.
При определении морфологических изменений следует учитывать влияние воздействие климатических и эдафических (почвенных) факторов на устойчивость растений. При газовом загрязнении на формирование некрозов решающим образом влияет влажность и освещенность. При высокой влажности воздуха и почвы растения становятся особенно восприимчивыми; зимой снижение устойчивости вызывается низкими температурами.
Очень большое экологическое значение и большую опасность для жизни человека, животных и растений имеют различные примеси к нормальному естественному составу воздуха. В связи с ростом промышленного производства в некоторых районах воздух перенасыщен газами и взвешенными частицами угля, металлической пыли, сажей и другими вредными примесями. Загрязнение воздуха вредными примесями в районах интенсивного производства неуклонно возрастает.
По данным Р. Гудериана (1979), выброс в атмосферу диоксида серы в ФРГс 1969 по 1980 г. увеличился с 3,5 до 4,6 млн. т; хлористого водорода с 1971 по 1980 г. — с 8 до 100 тыс. т. В США за период с 1960 по 1980 г. предполагается двукратное увеличение выбросов диоксида серы.
Во многих крупнейших городах, особенно в Токио, Лос-Анжелесе, Лондоне, Эссене и некоторых других, трудно дышать из-за смога — густого тумана, образованного выбросами труб промышленных предприятий, не снабженных очистными сооружениями, выхлопными газами автомобилей. Особенно вреден фотохимический смог, возникающий в результате взаимодействия дыма и выхлопных газов с солнечным излучением и содержащий большое количество оксида углерода. Выхлопные газы автомобильного транспорта наносят большой вред здоровью человека, так как в их составе имеются оксиды свинца, оксид углерода, оксиды азота и др.
Очень опасно заражение осадков радиоактивными веществами, которые способны вместе с воздушными массами дважды обогнуть земной шар, прежде чем выпасть на Землю. Необходимо принять самые действенные и неотложные меры борьбы с загрязнением воздуха вредными примесями. В числе этих мер кроме установки дымо- и газоулавливающих фильтров на заводских трубах и постройки других технических сооружений, которые не полностью очищают дым от газов и вредных примесей, необходимо обратить особое внимание на охрану и. восстановление нарушенного растительного покрова, являющегося естественным фильтром, очищающим 100 воздух от пыли и других вредных отходов производства. По данным Т. Келлера (Т. Keller, 1971), еловый лес, растущий на площади 1 га, способен связать 30 т, а буковый — 68 т пыли. Но задержанная на кронах деревьев пыль, в частности зола, образующаяся при сгорании бурого угля и содержащаяся в пыли, затем смывается дождем с поверхности растений и изменяет рН почвы. Это отрицательно сказывается на жизнедеятельности корней древесных растений. Газообразные соединения и соли, выделенные из пыли и принесенные талыми водами из золоотвалов, угнетающе действуют на естественную культурную растительность. Из культурных растений особенно неустойчив к ним картофель. Наиболее вынослив к избытку хлоридов овес, а из сорняков — пырей.
Растительность может задерживать пыль и обезвреживать газы до определенных пределов. Чрезмерное их количество сначала угнетает, а затем приводит к гибели малоустойчивые древесные и травянистые растения. Наиболее сильно растения повреждаются вредными отходами производства там, где их концентрация наибольшая: вблизи химических заводов, доменных печей, нефтеперегонных заводов и других загрязняющих воздух предприятий. При этом гибнут не только отдельные растения, но и леса, особенно хвойные. По мере удаления от источника загрязнения концентрация газов в воздухе снижается, и их отрицательное воздействие на растительность уменьшается. Однако неблагоприятное влияние кислых газов прослеживается на расстоянии до 5-7 км и даже до 30 км от источника задымления! В настоящее время многочисленными исследованиями показано, что наиболее вредное действие на растительность оказывает диоксид серы S02. Концентрация его в воздухе, равная одной миллионной доле и даже меньше от общего объема воздуха, значительно повреждает растительный покров. S02 вместе с другими газами (хлором, триоксидом серы, оксидами азота, углерода и т.д.) выделяется при топке печей каменным углем.
Сущность вредного влияния S02 на растения сводится к следующему; проникая в клетки тканей листа, он растворяется в воде и превращается в сернистую» кислоту, которая, накапливаясь в клеточном соке, вызывает повышенное окисление содержимого клеток. В окислении клеточного содержимого кроме S02 участвуют и другие кислые газы: хлор, триоксид серы, оксиды азота. При действии повышенных концентраций кислых газов, повышающих окисляемость клеточной среды, повреждаются хлоропласты и прилегающая к ним цитоплазма. Разрушаются хлорофилл, особенно хлорофилл «b», глютатион, аскорбиновая кислота и другие вещества клетки. Слабеет или приостанавливается фотосинтез, что приводит к уменьшению или прекращению выработки углеводов и других веществ, к задержке роста. Диоксид серы — сильный восстановитель, он изменяет и нарушает окислительно-восстановительное равновесие клеточной среды, уменьшает устойчивость биоколлоидов клетки.
Не все растения газоустойчивы в одинаковой степени. Например, хвойные — сосна и ель — повреждаются малыми концентрациями газов; другие переносят значительные концентрации S02 и других кислых газов без видимых повреждений. Если большие концентрации токсических газов воздуха губительны для растений, то малые и даже средние их величины не оказывают столь вредного влияния. В последнее время появляются работы, где показана приспособленность некоторых растений к газам и веществам, загрязняющим воздух. Накопились данные о том, что примеси к естественному составу воздуха даже стимулируют рост некоторых растений.
По мнению некоторых зарубежных ученых, стимулирующее влияние веществ, загрязняющих воздух, на рост объясняется тем, что некоторые растения, не получающие достаточного количества необходимых для их роста азотных соединений из почвы, используют диоксид азота из загрязненной атмосферы. По мере загрязнения воздуха растения постепенно приспосабливаются к нему. Многие исследователи считают, что необходимо создать особый раздел экологии — индустриальную экологию, который изучал бы адаптации растений к токсическим газам.
Нельзя смешивать понятия газоустойчивость и газочувствительность. Под газоустойчивостью понимают способность растений благодаря ана-томо-морфологическим, физиологическим и биохимическим особенностям выдерживать значительные концентрации токсических газов, сохраняя при этом свою жизненность и декоративность. Газочувствительность — это степень и быстрота проявления у растений признаков повреждения токсическими газами. В.М. Рябишга (1965) показал, что ель обыкновенная малочувствительна к S02 и другим кислым газам, но вместе с тем крайне неустойчива к ним, в то время как лиственница очень чувствительная и в тоже время довольно газоустойчивая порода. Газоустойчивость лиственницы объясняется тем, что она ежегодно обновляет свою хвою, в то время как хвоя ели многолетняя и подвергается воздействию токсических газов в течение ряда лет.
Н.П. Красинский (19S0), Е.И. Князева (1950) и другие авторы различают три вида газоустойчивости растений.
1. Биологическая газоустойчивость, под которой понимают способность растений быстро восстанавливать поврежденные дымовыми газами части и органы, а также декоративность. Например, клен американский (Acer ne—gundo) повреждается S02 средне, но в силу свойственной ему биологической активности очень быстро восстанавливается.
2. Морфолого-анатомическая газоустойчивость связана с морфологическими и анатомическими особенностями строения. Жесткие кожистые листья склерофитов, покрытые толстым слоем кутикулы, многослойным эпидермисом и восковым налетом, имеющие толстые наружные стенки клеток и другие признаки ксероморфной структуры,препятствующие проникновению токсических газов в цитоплазму к хлоропластам, менее повреждаются S02 и другими газами, чем листья тонкие, нежные, с тонкой кожицей, не покрытые восковым налетом. Так, ель обыкновенная отличается крайне слабой газоустойчивостью. Ока сильно угнетается при незначительной концентрации S02 в атмосфере. Однако американские ели — колючая (Picea pun—gens) и Энгельмана (P. engelmanm), особенно их голубые и серебристые формы, довольно устойчивы к этому газу. Хвоя этих елей покрыта восковым налетом, препятствующим проникновению оксида серы внутрь хвои -к клеткам мезофилла. Значительная газоустойчивость суккулентов также объясняется тем, что они имеют толстый слой покровных тканей, развитый восковой налет, малое количество устьиц. Все это, препятствуя проветриванию тканей листа, приводит к резкому снижению газообмена и к уменьшению проникновения токсических газов в клетки.
3. Физиологическая газоустойчивость зависит от физиолого-биохими-ческих особенностей растений. Некоторые представители семейства капустных, например, не способны к быстрому восстановлению поврежденных листьев и побегов и не имеют ясно выраженной ксероморфной структуры и других приспособлений, препятствующих проникновению газов в клетки листа, и тем не менее газоустойчивы. Их газоустойчивость можно объяснить только физиолого-биохимшчеекими свойствами цитоплазмы их клеток.
У большинства растений эти три вида газоустойчивости существуют одновременно, у других преобладает тот или иной вид гаэоустойчивости.
Загрязненность воздуха оказывает на растения угнетающее воздействие. Тормозятся ростовые процессы, значительно изменяется развитие растений: сдвигается цветение, сокращается вегетационный период, наблюдаются преждевременные листопад, изменение других фенофаз. Из всех органов растений в первую очередьи в наибольшей степени повреждаются газами листья.
Различают острые и хронические повреждения растений газами.
Острое повреждение происходит при действии на растения токсического вещества высокой концентрации в течение нескольких минут и часов и вызывает необратимые нарушения в структуре ассимиляционных тканей. Эти нарушения отчетливо различимы визуально по появлению ожогов, снижению тургора листьев, пониканию и в дальнейшем опаданию.
Хроническое повреждение растений происходит при систематическом проникновении в листья и стебли небольших доз токсикантов. Внешне симптомы выражены слабее. Характерными признаками хронического поражения растений являются сокращение плотности облиствения крон деревьев, уменьшение линейных размеров ассимиляционных органов, преждевременный листопад, снижение прироста деревьев по высоте и диаметру, частичная или полная потеря плодоношения, а в отдельных случаях формирование нежизнеспособных семян.
Степень повреждения оценивается по доле некротических участков в общей площади места (%) и по пятибальной системе со следующими значениями некроза листьев:
-
очень слабые ожоги листьев
-
слабые
-
средние
-
сильные
-
очень сильные
Степень повреждения растений кислыми газами зависит от влажности воздуха, температуры и других экологических факторов. В.Г. Актинов (1962), В.М. Рябинин (1965) и другие исследователи показали, что в местах с повышенной влажностью воздуха дым и газы концентрируются в нижних слоях атмосферы. По этой причине в низинах, где обычно застаиваются туманы, наблюдается острое отравление растений диоксидом и трн-оксидом серы. Повреждения растений токсическими газами особенно усиливаются при одновременном повышении относительной влажности и температуры воздуха. Концентрация S02 и других газов в воздухе зависит от силы и направления ветра по отношению к источнику задымления. Как правило, с повышением скорости ветра, особенно на открытых местах, концентрация газов в приземном слое воздуха понижается.
На газоустойчивость растений влияет режим питания, особенно азотного. В.А. Гусева (1950) показала, что при подкормке растений нитратной формой азотных удобрений (NaN03) общая окисляемое» клеточного содержимого окуренных S02 растений и окисляемое» водонерастворимых веществ снижаются, уменьшается разрушение аскорбиновой кислоты и глютатиона.Впоследствие этого газоустойчивость подкормленных селитрой растений по сравнению с контрольными значительно повышается. Однако аммонийная соль действует на газоустойчивость значительно слабее селитры.
Газоустойчивость разных групп растений далеко не одинакова. Особенно чувствительны к S02 и другим газам лишайники, мхи и некоторые эпифиты. Наствольные лишайники — наиболее надежные показатели чистого воздуха, поэтому они отсутствуют на деревьях в промышленных городах. Неустойчивость лишайников к загрязнению атмосферы дымовыми газами Р. Гудериан (1979) объясняет их слабой регенеративной способностью. В то время как у высших растений, особенно у однолетников, поврежденные газами ассимиляционные ткани быстро заменяются новыми, чаще всего менее чувствительными к дымовым газам, ткани лишайников растут очень медленно, и в условиях долговременного воздействия газов их отравление продолжается до полной гибели всего слоевища. Лишайники особенно чувствительны к дымовым газам при длительном дефиците влаги. Травянистые растения, как правило, более устойчивы к дымовым газам, чем древесные.
Повреждаемость деревьев и кустарников токсическими газами также не одинакова. Особенно слабоустойчивы к дымовым газам сосна и ель. H.JI. Красинский показал, что газоустойчивость растений определяется их систематическим положением. Наиболее газоустойчивы растения из семейств ивовые (Salicaceae) и жимолостные (Caprifoliaceae), среднеус-тойчивы представители кленовых (Асегасеае), камнеломковых (Saxi-fragaceae) и маслинных (Oleaceae), очень сильно повреждаются кислыми газами виды семейства бобовых (Leguminosae).
2.2 Патологические проявления у животных
Язвы на коже
Язвы на коже, наблюдались у многих видов рыб, отловленных вблизи побережья и вдали от него. У трески обитающей в водах Северной Европы, частое появление этих язв получило название “ язвенного синдрома”. Недавнее исследования продемонстрировали связь между сезонными изменениями этого синдрома, степень о загрязнения вод углеводородами и увеличением в воде популяций, потенциально патогенных для рыб. Весной процент больных рыб у побережья выше,чем вдали, но к лету эта зависимость становится менее очевидной. Поэтому при отлове рыб с целью мониторинга нужно учитывать сезон года. Кроме того, следует проводить микробиологические тесты проб, взятых из донных осадков и водной толщи.
Эрозия плавников
Это одно из наиболее распространенных заболеваний рыб, четко связанное с загрязнением эстуарной и прибрежной среды.
Встречаются два типа эрозии плавников. У приданных рыб, видимо, в результате прямого контакта с загрязненными донными осадками поражаются спинной и. анальный плавники, a у пелагических прибрежных рыб наблюдается общая эрозия, но с некоторым преимущественным поражением хвостового плавника. Вероятней всего, причины эрозии носят комплексный характер и могут включать химическиё агенты (которые влияют на мускус эпителий), дефицит растворенного кислорода в воде и вторичное бактериальное заражение. Систематическое заражение бактериями не обязательно связано с появлением эрозии плавика, хотя в пробе, взятой из язвы, можно выделить многие виды бактерий. Наблюдение за этим показателем рекомендуется проводить с учетом сезона года, размера рыб, чувствительности вида, условий обитания и миграции.
Аномалии скелета
3a последние годы увеличилось число случаев аномалии скелета у рыб. Приводится множество примеров спинных фузий и искривлений, позвоночного сжатия (уплощение), аномалий головы и плавника. Такие нарушения встречаются и у большинства природных популяций, no чаще всего они наблюдаются в загрязненных акваториях. Связь между частотой появления аномалий скелета водных позвоночных и загрязнением была подтверждена экспериментально. Хлорорганический пестицид Кепон, например, вызывал сколиоз у миног, при действии тяжелых металлов у рыб наблюдались искривления и разрывы позвоночника. Таким образом, мониторинг морских позвоночных на тканевом уровне включает тщательный осмотр рыб для выявления явных аномалий с последующей рентгеноскопией для обнаружения скрытых деформаций, например позвоночныхспаек. He представляет особых трудностей обследование жаберных тычинок и спинных плавников. Большую пользу могут оказать планктонные и планктоновые пробы с целью обнаружения уродливых личинок и аномалий у ранней молоди.
Опухоли
Опухоли были обнаружены у представителей всех классов холоднокровных позвоночных, у двустворчатых моллюсков и насекомых. У 60 морских видов животных из разных групп и местообитаний были обнаружены инфекционные опухоли. Опухоли у рыб и моллюсков являются потенциально полезными показателям для мониторинга морской среды, по ограниченное географическое распространение видов, имеющих опухоли, и отсутствие опухолей у видов, имеющих широкое географическое распространение, a также недостаток данных о причинах; вызывающих опухоли, исключают возможность использования какого-то одного вида морской рыбы в качестве универсального индикатора.
Использование двустворчатых моллюсков для мониторинга химических карциногенов в окружающей среде имеет значительные преимущества, так как они, в отличие от рыб, например, профильтровывают большие количества воды в течение длительного времени. Важно также то,что относительно небольшое число видов обитает почти во всех эстуариях Мирового океана.
Более того, рак крови у двустворок описан для четырех континентов, и, хотя имеются доказательства в пользу как вирусной, так и химической этиологии этого заболевания, его появление,по-видимому, связано с присутствием загрязняющих Методика определения рака крови у моллюсков чрезвычайно проста и состоит в наблюдении за мутностью 0,5 смз жидкости, полученной из тела. Нормальные гемоциты прилипают стеклу, и капля быстро становится прозрачной. Раковые клетки становятся круглыми, не прилипают к стеклу, и сама капля очень похожа на каплю молока.
Иммунная реакция
Современными иммунологическими исследованиями показано, что рыбы в противоположность беспозвоночным вырабатывают высоко специфические антитела. Оказалось, что антитела, патогенные для человека, встречаются в 1,5-6% рыб мороне обитающих в заливе Чесапик. Это было отмечено преимущественно в эстуарных районах вблизи крупных поселений, причем кросскорреляция среди антител небольшая.
Имеется очень мало доказательств подавления иммунных реакций непосредственно загрязнениями. Так, при действии кадмия на рыб обнаружено снижение эффективности реакции фагоцитов. Были получены данные о генетическом отборе рыб с более высокими уровнями антител.
Таким образом, тест на присутствие у рыб антител, патогенных для человека, может стать важной частью мониторинга, особенно ценной для тех видов рыб, которые имеют строго ограниченный ареал обитания.
Лимфоцитоз
Лимфоцитоз — заболевание, широко распространенное среди плоских рыб. Оно характеризуется образованием опухолеподобной ткани под кожей, содержащей гипертрофированные фибробласты размером до 2 мм. Это заболевание имеет вирусное происхождение. B некоторых случаях они могут быть переносчиками паразитов рыб. Имеются данные о том, что лимфоцитозом чаще заболевают рыбы, обитающие в загрязненных и нагретых водах. Поэтому этот показатель можно использовать для мониторинга, хотя следует иметь в виду, что лимфоцитоз иногда встречается среди популяций рыб и в незагрязненном районе в результате эпидемии. Известно, что некоторые болезни рыб находятся в скрытом состоянии и могут быть спровоцированы при действии неблагоприятных факторов. таким образом, лимфоцитоз служит ранним сигналом неблагополучного состояния среды и может в определенных условиях успешно использоваться в системах мониторинга.
Жаберная гиперплазия
Рыба может реагировать на химические вещества гиперплазией (клеточной пролиферацией)жаберного эпителия. Но количественная связь “причина-эффект” не известна, и ее еще предстоит выявить. Получить жаберные пробы не представляет особого труда.
Изменение ткани печени
Известно, что печень играет важную роль в детоксикации выведении загрязнении. Имеются экспериментальные данные том, что под действием загрязнения происходят различные паралогические изменения в ткани печени, в том числе образование, опухолей
3. Организменный уровень
Основная цель отбора биологических показателей на организменном уровне сводится к оценке главным образом физиологического состояния особи и ее поведения, a также биологической значимости данных по концентрации загрязняющюших веществ в теле организмов. Однако эти данные будут представлять интерес только в том случае, если изменения величины отклика используемых показателей коррелируют с изменениями на экосистемном уровне, т, е. если изменения физиологических показателей приведут к изменению экологической слаженности.
Таким образом, при биоиндикации на организменном уровне используются следующие признаки:
1. изменение окраски листьев и тела животных, а также скульптуры поверхности
2. изменение размеров и продуктивности растений и животных;
3. изменение формы роста, экобиоморфных признаков;
4. показатели поведения животных.
5. биоритмические изменения – основными факторами, определяющими цикличность, выступают освещенность, температура, влажность;
3.1 Изменение окраски листьев и тела животных
а) изменение окраски листьев — в большинстве случаев неспецифическая реакция на стрессоры:
— хлороз — бледная окраска листьев между жилками (тяжелые металлы, соединения хлора,хлор); хлороз вызывается нарушением соотношения в организме Fe и Са. В естественных условиях он может возникать у растений при нарушениях водного, светового режима, оглеении и заболачивании почв. Встречаются межжилковый, верхушечный и другие виды хлороза. Степень повреждения листьев может быть выражена в балльной системе и использоваться как количественная характеристика при анализе и картировании распространения газообразных токсикантов.
— пожелтение листьев или определенных их участков (у лиственных пород под влиянием хлоридов);
— покраснение листьев — накопление пигмента антоциана под действием SO2;
— побурение и побронзовение (начальная стадия тяжелых некротических повреждений, под действием О3);
— окраска, при которой листья имеют вид пропитанных водой, сходная с морозными поврежедениями (начальная стадия некрозов, вызванных пероксиацетилнитратом);
б) изменения окраски тела животных. Среди реакций окраски тела на антропогенное изменение среды, наиболее известной является “индустриальный меланизм” или “меланизм крупных городов”. Под меланизмом подразумевается заметное потемнение первоначально светлых форм.
в) Изменения скульптуры поверхности. Установлена четкая корреляция между структурой поверхности тела, которая изучается с помощью электронного микроскопа и уровнем загрязнения среды — объектами исследования также являются насекомые.
3.2 Изменение размеров и продуктивности растений и животных
а) изменения размеров органов растений и их положения — чаще всего неспецифичны, так хвоя укорачивается под действием SO2 и удлиняется под действием нитратов; аномальная конфигурация листьев отмечается у лиственных деревьев после радиоактивного облучения; в результате локальных некрозов возникает уродливая деформация, перетягивание, вздувание одельных участков листовой пластинки; искривление побегов; сращение и расщепление отдельных органов; увеличение или уменьшение частей цветка;
б) изменение прироста и плодовитости растений — чаще всего неспецифичны, однако широко применяются для биоиндикации; измеряют в основном радиальный прирост древесных стволов,прирост в длину побегов и листьев, длину корней; уменьшение образования плодовых тел у грибов, продуктивности у черники и елей в районах, загрязненных газообразными выбросами.
в) изменение структуры древесины — снижение качества сосновой древесины в результате незначительного ее образования летом и выпадения годичных колец под действием SO2; одревеснение корней злаков при обработке гербицидами.
Кресс-салат. Рекомендуется для индикации хлоридов и свинцовых солей и загрязнения воздуха выхлопными газами. В качестве показателей используют длину проростков и корешков, число семян.
Липа как индикатор соли, применяемой для таяния льда на улицах. Во многих европейских странах применяется индикация по изменению окраски листьев липы: сначала появляются ярко желтые краевые зоны, они продвигаются к середине и к основанию листа.
Измерения прироста хвойных пород, кустарников разнотравья и злаков. Наиболее известным методом является измерение годичных колец у деревьев. Для установление ширины годичного кольца у пробных деревьев отбираются радиальные буровые керны; затем рентгеновским денситометром определяют плотность древесины, т.е ее качество. Снижение объемного прироста на территориях с сильным загрязнением SO2, засолением почвы или значительной рекреационной нагрузкой составляет у хвойных около 50%.
У кустарников продуктивность определяется измерением годичных побегов, а у разнотравья и злаков — суммы длин листьев.
г) Биометрические параметры – размеры тела и отдельных его частей у животных. Изменения размера животных можно учесть, используя уравнение аллометрического роста v=аMb (где v -скорость роста, M — масса тела, a – постоянная b — показатель). .
Для биоиндикации антропогенных стрессоров привлекаются биометрические параметры главным образом различных насекомых — промеры тела: ширины головы, длины различных члеников, усиков, конечностей. Использование этих биометрических параметров для индикации антропогенных стрессоров сталкивается с одинаковыми проблемами — даже при статистически значимых отличиях, трудно доказать их связь с градиентом рассматриваемых нагрузок, обусловленных человеческой деятельностью.
3.3 Изменение формы роста и экобиоморфных признаков
Измерения физиологических показателей можно приурочить к какой-то фазе репродуктивного цикла или к какому-то времени года. Реакция организмов на загрязнение не всегда линейно за висит от действующей концентрации, так как существует пороговый уровень, ниже которого организмы могут детоксифицировать вещество. Аномальные отклики сигнализируют о том, что этот порог превышен. При надпороговых значениях концентрации вещества физиологическая реакция будет соответствовать величине действующего фактора. Если перенести организм в чистую среду, то он снова окажется способным к детоксикации загрязнения, что, в конечном счете, приведет к нивелированию отрицательного отклика
Рост.
Рост – один из важнейших параметров, характеризующих состояние популяции в конкретных условиях окружающей среды. Поскольку снижение скорости роста животных является неспецифическим откликом на любые стрессовые воздействия, следует быть очень внимательными при использовании этого показателя и интерпретации получаемых данных. 3амедление роста может наступить из-за снижения интенсивности питания или увеличения расхода энергии, связанного с дыханием или выделением, под действием разных экологических факторов – температуры, солености, концентрации 02, концентрации пищи, ионов металлов, нефти, скорости движения воды и др. Рост отдельных особей можно измерять прямым методом или косвенно. Непрямой метод, или физиологическая оценка роста является интегральным, поскольку включает измерение таких физиологических переменных, которые отдельно и вместе характеризуют состояние особи.
Несмотря на то, что лабораторные и полевые исследования по влиянию загрязнения на показатели роста в основном выполнены на ракообразных и двустворчатых моллюсках, a для рыб имеются данные только лабораторных опытов, этот показатель следует активно использовать в программах мониторинга.
Питание.
Для животных характерны разнообразные способ питания травоядность, питание детритом и седиментами фильтрация, некрофагия и хищничество. Процесс питания сложный и включает нетолько непосредственно пищеварение, но и поведенческие компоненты: поиск пищи, выбор и поимку жертвы, фильтрацию пищи из водного столба, добывание в донных осадках и др. Изменение скорости питания и образование фекалий служат показателями физиологического состояния в условиях загрязнения. Показатель скорости питание в естественных условиях изменяется в широких пределах. Он весьма чувствителен даже к небольшим изменениям количества доступной пищи, что сильно усложняет задачу определения фонового уровня.
Дыхание.
Скорость потребления кислорода. По этому показателю можно судить о физиологическом состоянии как целого организма, а так и изолированных тканей.Однако интерпретация данных измерений по потреблению кислорода затруднена из-за того, что обычно не учитывается влияние других экологических факторов, таких, как соленость, температура, время года и др. Респираторная активность. B программах мониторинга используются три показателя активности дыхания главным образом у пресноводных рыб: частота кашля, частота оперкулярного движения и частота сердцебиений. Частота кашля изменяется под действием физических (например, взвешенные твердые частицы) или химических (например, тяжелые металлы) факторовна эпителий жабр. Измерения частоты оперкулярного движения проводились на многих видах рыб в присутствии разных загрязнителей изменение обычно связано с уровнем потребления кислорода, хотя последний может быть сбалансирован за счет регуляции амплитуды движений. Изменение частоты сердцебиения также связано с изменениями потребления кислорода, хотя и в этом случае компенсация может быть достигнута путем изменения глубины дыхания. Из трех показателей частота кашля,по-видимому, наиболее чувствительный и удобный для исследователя отклик на широкий круг загрязнений.
Выделение и азотный баланс
Продукты выделения морских животных включают аммиак, мочу, аминокислоты и пурины.
Баланс между поступающими в организм веществами с пищей и их выделением с продуктами жизнедеятельности обычно рассчитывают с помощью уравнения энергетического баланса и показателей роста. В тех случаях, когда содержание азота в пище лимитировано, потеря азота в выделениях может свидетельствовать об азотном нарушении баланса. Скорость выделения, превышающая значения нормы, может свидетельствовать о стрессовой ситуации, вызванной серьезным нарушением питания под влиянием загрязнения. Скорость выделения азота может дать больше информации о состоянии животного, если ее рассматривать наряду с другим физиологическими показателями. Отношение потребленного кислорода к выделенному азоту (отношение O/N) является индексом катаболического баланса белка, углеводов и липоидов, там как атомные эквиваленты потребленного кислорода при катабализме и выдeленного азота колеблются. Высокое значение индекса O/N указывает на преобладание липидного или угле водного катаболизма над распадом белков. Теоретическое минимальное значение O/N при исключительно белковом катя холизме равно приблизительно 7.
Воспроизводство.
Снижение значений показателей роста может привести к снижению плодовитости животного, так как на образование гамет расходуется существенная часть запасаемой энергии. Любые наблюдаемые снижения показателей роста, особенно во время созревания гамет, могут быть сигналом резкого снижение репродуктивной способности родительского организма. Эмбриональные и личиночные стадии развития, как правило, наиболее чувствительны к токсикантам по сравнению со взрослыми особями. При изучении действия загрязняющего вещества на воспроизводство следует иметь в виду, что часть его может накапливаться в икринках и сохраняться там длительное время. Контроль за этим накоплением позволяет с некоторой долей вероятности прогнозировать последующие стадии развития организма.
Определение плодовитости и оценка жизнеспособности гамет являются полезными показателями состояния особей, с помощью которых можно объяснить некоторые экологические нарушения.
Состав крови.
При оценке состояния рыб и их откликов на изменение окружающей среды в последние годы стали чаще использовать гематологические методы. Исследования крови беспозвоночных в связи с влиянием фактора окружающей среды немногочисленны.
3.4 Показатели поведения животных
Первый этап на пути использования показателей поведения в программах мониторинга морской среды заключается в разработке качественных и количественных критериев оценки изменения поведения при действии антропогенных факторов. Различают три подхода для оценки поведенческих реакций. Первый подход предполагает наблюдение за поведением животных, обитающих в естественных условиях. Второй — предусматривает перенос животных из лабораторных условий или контролируемых акваторий на какое-то время в естественные условия. Третий подход заключается в переносе воды и донных осадков из естественных условий в аквариумы, в которые помещают тест организмы. Окончательное решение вопроса о включении показателей поведения в программы мониторинга принимают после оценки их экологической значимости и связи с уровнем накопления в окружающей среде и контролируемых организмах.
Выбор организма или групп организмов — первый, необходимый шаг при разработке программ биологического мониторинга — основывается, прежде всего, на экосистеме, которая подвергается антропогенному воздействию, Выбранный для мониторинга организм должен при появлении загрязнения своим поведением сигнализировать об изменившихся условиях. Чем чувствительнее поведенческие отклики к появлению химических вещeств и чем теснее они связаны с целостными свойствами экосистем, тем эффективнее использование выбранных организмов. Поведенческий отклик y любого вида формируется под влиянием разнообразных стимyлов. Но все они,так или иначе, связаны с общей стратегией поведения вида, направленной на выжигание. У морских организмов структура поведенческих реакций может видоизменяться в ходе индивидуального развития. Поэтому конкретная программа мониторинга должна учитывать этологическое особенности разных стадий развития организма, a также изменения этик показателей под влиянием сезонных, суточных и многолетних циклов. Однако если по каким-либо причинам этот поведенческий инстинкт нарушить, это приведет к снижению численности популяции из-за их потребления хищниками или гибели в холодной воде.
Для обнаружения изменений качества морской среды в определенной акватории выбирают контрольную станцию, в которой диапазон изменчивости модели поведения соответствует норме.
Отклонения от нормы в поведении у животных, обитающих в районе загрязнения, свидетельствуют об изменении качества морской среды. Значительные изменения структуры всего сообщества сопровождаются, как правило, выпадением из сообщества одного или нескольких видов. Изменение модели поведения может вызвать также нарушение трофических связен. Haпpимep, необратимые нарушения хеморесценции или локомоции постепенно приведут к резким изменениям трофических отношений. Изменение привычного поведения хищника тут же отражается на изменении показателей популяции жертвы при условии, если жертва менее чувствительна к загрязнению, чем хищник. Если модель поведения изменяется под действием токсиканта, то это заведомо приводит,к изменению выживаемости вида, a в случае, если это доминантный вид, — ас существенным структурным изменениям в сообществе.
Выживаемость организмов в морской среде обеспечивается за счет комплекса сложных поведенческих реакций, которые сформировались в ходе длительной эволюции организмов. Чувствительность и адаптация к изменениям в окружающей среде, способы захвата пищи, избежание хищников и воспроизводство неразрывно связаны с условиями окружающей среды. Поэтому, например, сенсорную чувствительность и связанные с ней другие разновидности поведенческих реакций можно использовать для оценки действия загрязнения, если они связаны c выживаемостью организма. Любое регистрируемое изменение сенсорной чувствительности или модели поведения,связанной с ней, будет показателем степени воздействия загрязняющиx веществ.
Морские организмы реагируют на присутствие в воде естественных и искусственных веществ и очень небольших концентрациях. Более сложные поведенческие реакции, такие, как поиск пищи или спаривание, меньше пригодны для программ мониторинга. Их использование оправдано только в том случае, если биология тест организма и особенно его поведенческая реакция достаточно хорошо изучены. Таким образом, причиной изменения в поведении животных под действием загрязняющих веществ могут быть как изменение чувствительности хеморецепторов, Так и общее изменение их ответной реакции. Для того чтобы установить причины изменения поведенческой реакции, необходимо проводить нейрофизиологический контроль состояния хеморецепторов.
Контрольные вопросы:
1. Критерии отбора биоиндикационных показателей тканевого и организменного уровня.
2. Показательные признаки поврежедения на ткневом уровне, примеры.
3. Характеристика и типы некрозов у растений.
4. Характеристика стандарнтных тест-растений для биоиндикации на тканевом уровне.
5. Показательные патологические проявления неблагоприятного внешнего воздействия у
животных как биоиндикацилонные показатели, примеры.
6. Показательные повреждения организменного уровня у растений, примеры.
7. Изменение окраски листьев у растений и тела у животных как биоиндикационные показатели, примеры.
8. Изменение размеров и продуктивности растений и животных как биоиндикационные признаки.
9. экобиоморфные признаки как биоиндикационные покажзатели, примеры.
Лекция 4
Биоиндикация на высших иерархических уровнях: популяция, экосистема, биоценоз
План
1. Популяционный уровень
Отбор показательных видов
Показатели популяционного уровня
Воздействие антропогенных стрессоров на динамику растительных популяций
Воздействие антропогенных стрессоров на характер распространения растений
2. Экосистемный уровень
Показательные признаки экосистемного уровня
Метод комплексной биоиндикации
1. Популяционный уровень
1.1 Отбор показательных видов
1 вопрос лекции. Отбор показательных видов
Для отбора видов необходимы подробные данные о биоте изучаемого района. Объектом наблюдения может быть любая группа организмов: от микрофлоры до мегафауны. B тех случаях, когда предусмотрено использование относительно малоизученных видов, обязательно исследование их физиологии. При отборе видов следует учитывать их пространственное распределение. Ограниченная доступность может вызвать необходимость многократного отбора проб, a это в свою очередь вынуждает разрабатывать соответствующие требования в отношении объема выборки и частоты их взятая. Предпочтение следует отдавать видам, чувствительным к потенциальным загрязнениям, даже если они имеют ограниченное экологическое и промысловое значение.
Трудности отбора проб, связаны с особенностями поведения организмов в зависимости от сезона, возраста и миграций. При разработке программ биоиндикации следует брать широко распространенные в изучаемом районе виды. Это позволяет увеличить число наблюдаемых станций и изучаемых популяций, доступных для исследования. Отбор относительно равномерно заселенных мест обитания выбранного вида является необходимой частью последующих шагов, связанных с разработкой методов отбора проб. Эти меры позволяют уменьшить влияние естественной измен-чивости — эту вечную проблему отбора проб в биологии. При отборе видов для биоиндикации следует отдавать предпочтение видам, представляющим различные трофические уровни.
1.2 Показатели популяционного уровня
Использование показателей популяционного уровня зависит от выбранных видов.
а) Ростовые показатели. Данные об абсолютной или относительной скорости роста можнополучить, изучая структуру популяции или видов известного возраста. Поскольку маркировку организмов проводить очень трудно, лучше всего использовать такие виды, у которых образуются ежегодные метки.
б) Изменение плодовитости особей, входящих в популяцию, может свидетельствовать о нарушении репродуктивного процесса. Этот показатель целесообразно использовать для видов, которые откладывают относительно небольшое число яиц.
в) Распределение и обилие видов. Распределение и обилие являются особо ценными показателям состояния популяции оседлых видов, особенно при изучении градиента загрязнения. Однако показатель обилия для подвижных форм обладает большой изменчивостью и низкой разрешающей способностью, поэтому его ценность для биоиндикации сравнительно невелика.
г) Структура популяции. Для оценки этого показателя можно использовать методы определения возрастных групп. Максимальная чувствительность структурного показателя популяции достигается при наблюдении за изменениями динамики популяции.
д) Биомасса — это структурный показатель, определение которого не представляет больших трудностей. В травянистых сообществах удобнее и проще всего проводить учет биомассы в период максимального развития травостоя. Одним из наиболее распространенных методов является метод укосов. Для этого на пробной площади размером 20х25 м размещают 20 площадок 0,25х0,25м, 0,5х0,5 м или 1х1м. Выбор оптимального размера площадок определяется общими запасами и богатством травостоя.
е) Обилие. Величина обилия меняется в больших пределах, чем биомасса, если она рассматривается для определенных размерных классов. Суммарное значение обилия, по-видимому,менее изменчиво, чем обилие отдельных популяций.
ж) Видовое разнообразие. Число видов в данном таксоцене обычно сильно зависят от числа и размера взятой пробы. Измерение разнообразия основано на суммарном числе видов и особей и относительном обилии особей данного вида.
з) Число высших таксонов. Простым методом биоиндикации может быть регистрация числа особей c определением их до рода, семейства или порядка. Например, обнаружено постепенное снижение числа таксономических групп мейофауны в направлении от открытого моря к загрязненным прибрежным зонам Южной бухты северного моря: на некоторых станциях оказалась всего одна группа (нематоды) или две (нематоды и копеподы) по сравнению c десятью группами на незагрязненных станциях прибрежных вод. Эти группы мейофауны легко определяются, и их число может служить в качестве полезного показателя для мониторинга антропогенных изменений.
и) Трофическая структура
Трофическое положение вида имеет важное значение для таких процессов, как биоаккумуляция и общий поток энергии в сообществе. Соотношение первичных продуцентов или консументов может быть связано c сукцессией и стабильностью сообщества, a соотношение видов c различными типами питания может указывать на преобладающий вид энергии, доступный сообществу.
к) Сравнение сообществ Методы сравнения сообществ в пространстве и времени по своей природе носят статистический характер. Многие методы сейчас широко используются и уже имеются в виде стандартных вычислительных программ.
1.3. Воздействие на динамику растительных популяций
Антропогенные стрессоры могут влиять на все признаки растительных популяций. Наиболее чувствительно реагирует продуктивность. Она может многократно возрасти в результате ослабления конкурирующих видов. Изменяются также рождаемость и смертность, а в результате плотность популяции (число особей/площадь). Популяции с малой численностью особей находятся под особенно большой угрозой. В ответ на антропогенное нарушение происходит расширение или сокращение ареала популяции. В крайнем случае это может привести к исчезновению популяции и к вымиранию вида.
Очень важен для биоиндикации отбор устойчивых экотипов к действию антропогенных стрессоров. Экотип — совокупность особей любого вида организмов, приспособленная к условиям местообитания и обладающая наследственными признаками, обусловленными экологически. В природе разнообразие экотипов служит основой для поддержания одинаковой продуктивности популяции при переменных условиях обитания. При этом популяции многих видов содержат экотипы с высокой толерантностью к определенным антропогенным стрессорам. Биоиндикационное значение имеет тот факт, что нередко в поразительно короткие сроки (один или несколько вегетационных периодов) происходит вытеснение чувствительных и распространение устойчивых экотипов. Например, известны случаи отбора экотипов, устойчивых к тяжелым металлам, солям и сернистому газу. При сравнении береговой и растущей в отдалении от берегов популяции Agrostis stolonifera первая оказалась значительно солеустойчивей. Мак-самосейка обладает значительной устойчивостью к гербицидам — он активно проникает в массивы зерновых, где раньше встречался только по окрайкам полей.
1.4 Воздействие антропогенных стрессоров на характер распространения растений
Изменение ареалов растений. Возросшая нагрузка на ландшафт (земледелие, выпас, инустриализация) вызвала во многих странах резкое сокращение лесных площадей, ареалов древесных пород и видов нижнего лесного яруса. Безлесные и слабооблесенные ландшафты благоприятствуют вселению (иммиграции) многочисленных однолетних и многолетних растений.
Изменение местообитаний дало возможность многим видам продвинуться из своих исходных областей распространения на открытые, характеризующиеся теплым летом. Поэтому окультуренные ландшафты характеризуются медленным нарастанием числа адвентивных видов. Адвентивные или дополнительные виды — это виды, растущие в нетипичных местах.
Обеднение флоры. Ускоряется в результате устранения некоторых особых местообитаний полевых меж и тропинок, откосов, природных каменных стенок, переувлажененных участков, перелесков, суходольных лугов.
Причины уменьшения числа видов дикорастущих растений:
* устранение особых местообитаний — 200 видов
* осушение — 180 видов
* землепользование — 180 видов
* чрезмерная обработка почвы — 160 видов
* разработка, выемка почвы — 120 видов
* механические воздействия:
вытаптывание, стоянки — 100 видов
– применение гербицидов — 90 видов
– прополка, корчевание, выжигание — 80 видов
– застаивание вод — 70 видов
– сбор — 70 видов
– загрязнение водоемов — 30 видов
– урбанизация деревень — 20 видов
Ареалы растительных видов сильно изменяются под действием антропогенного стресса.
Это выражается в сокращении или расширении ареала различных растений.
Например, распространение накипного лишайника — относительно дымоустойчивого вида — позволяет оценить интенсивность многолетнего загрязнения воздуха данной территории.
2. Экосистемный уровень
2.1 Показательные признаки экосистемного уровня
Негативное воздействие анторопогенных факторов на природные экосистемы проявляется в изменении их видового состава. Обладая определенной устойчивостью к негативным воздействиям, виды формируют сообщества, отличающиеся биоразнообразием, которое может быть использовано для биоиндикационных целей. Показательными признаками являются:
1) видовой состав, видовое разнообразие;
2) характер распространения вида; динамика ареала, в том числе расширение ареалов синантропных видов;
3) популяционный анализ (продуктивность, плотность, динамика ареала, возрастная структура, смертность, внутривидовая дифференцировка, отбор устойчивых экотипов).
2.2 Метод комплексной биоиндикации
Оценка воздействия загрязняющих веществ на экосистемном уровне сводится к использованию данных, полученых для уровней популяции или сообществ, из которых оно состоит. Структурной основой экосистемы являются неорганические и органические вещества, факторы среды(температура, свет, ветер и др.), продуценты, консументы и, редуценты. Сложные взаимозависимые процессы функционирования экосистемы осуществляются за счет потока энергии, пищевых цепей, круговорота питательных веществ, изменения разнообразия, развития и эволюции во времени и пространстве, продуктивным оказалось предварительное выяснение чувствительных звеньев экосистем, по которым можно судить о состоянии экосистем.
Для оценки состояния окружающей среды используется метод комплексной биоиндикации.Он заключается в комбинации фито-и зооиндикаторов на разных системных уровнях.
Преимуществом этого метода является:
— возможность оценить состояние экосистемы как результат взаимодействия природных элементов экосистем и антропогенного воздействия,
— выявить степень устойчивости и реакцию экосистем на воздействие человека;
— использование биоиндикаторов, а именно их физиологических индикационных признаков, позволяет определить изменения в экосистемах на очень ранних стадиях, когда они еще не проявляются морфологическими и структурными изменениями и их нельзя выявить другими методами;
— возможность предвидеть нарушения экосистем и вовремя принимать меры, чтобы защитить экосистему от ущерба и тем самым не допустить больших народнохозяйственных потерь.
Методика имеет несколько последовательных этапов, на которых используются разные методы (рисунок).
Во-первых: выявление структуры территории по физико-географическим и экономико-географическим факторам, выявление антропогенного воздействия и на основе этого — определение однородных географических единиц.
Во-вторых: определение сети ключевых участков и их характеристик. Для равных типов экосистем необходимо иметь несколько однотипных ключевых участков, которые отличаются лишь степенью антропогенного воздействия.
Рисунок — Логическая схема исследований по методу комплексной биоиндикации
В — третьих: выбор индикаторов и биоиндикационные исследования выбранных индикаторов, проводятся в течение трех лет три раза в год — весной, летом и осенью, чтобы выявить не только состояние индикаторов, но и динамику их изменений.
В-четвертых: сбор дополнительной информации о состоянии почвы, воды и хозяйственной деятельности человека.
В-пятых: оценка состояния окружающей среды, которое имеет два этатпа:
1) выявление основных количественных соотношений между состоянием индикаторов исостоянием экосистем; построение оценочных шкал;
2) экстраполяция состояний экосистем на более крупные однородные географические единицы. Построение карты состояния окружающей среды. Биоиндикационные наблюдения ведутся на двух системных уровнях — популяции и индивида. Для каждого уровня разработаны критерии для выбора индикаторов и индикационных признаков, которые сведены в таблицу.
Степень изменения экосистем отражается экологическими индексами этих родов.
Биомасса сообществ оказалась лучшей характеристикой по сравнению с численностью, так как иногда освобожденную экологическую нишу после выпадения крупных малочисленных видов жуков занимают мелкие многочисленные виды.
На уровне индивидуума выбирают индикационные признаки по следующим критериям: тесная связь со значимыми для жизни реакциями организма; способность отражать постепенные изменения состояния организма, причем в ранних стадиях, пока они не проявляются в патологическом состоянии организма; способность отражать разную силу и качество фактора, возбуждающего изменения, вернуться в первоначальное состояние в связи с адаптацией организма к фактору или с исчезновением фактора; стандартный и простой способ отбора проб и их сохранения, стандартные и простые методы анализа.
Последние два критерия особенно важны при физиологических наблюдениях, так как большинство из них нуждаются в соблюдении точных экспериментальных условий.
На уровне индивида мы выбрали фито- и зооиндикаторы и следующие индикационные признаки.
Основной критерий, по которому определяется состояние фитоиндикатора — уровень фотосинтеза. Фотосинтез определяет жизнеспособность растений, их урожайность и хорошо отражает все изменения среды обитания. Но его уровень очень трудно измерить непосредственно, поэтому используются косвенные индикационные признаки — сухая биомасса, количество растворимых белков, активность фермента пероксидаза. Чем сильнее меняется состояние экосистемы в нежелательном направлении, тем меньше становится фотосинтез. В зеленых наземных частях растений уменьшается их сухая биомасса, концентрация растворимых белков и активность фотосинтетических ферментов.
Зооиндикаторы: грызуны родов Microtus и Apodemus. Эти два рода часто встречаются на изучаемой территории и их экологические особенности хорошо изучены. Индикационные признаки — масса организма, масса его органов (печени, почек), содержание тяжелых металлов в органах и шерсти, активность фермента церулопласмина.
Церулопласмин является одним из стрессовых ферментов. Это фермент крови, который содержит в своей молекуле ионы Cu (он голубой) и четко отражает состояние организма, повышая свою активность при раке, воспалениях и других патологических состояниях. Мы показали, что он также четко отражает и воздействие факторов среды обитания, например, при введении в организм тяжелых металлов и при иммобилизации животного. Это повышение соответствует силе воздействия — концентрации металла.
Состояние экосистем мы определяли сопоставлением состояния индикаторов – отклонением их индикационных признаков от фонового (оптимального) состояния. Для построения оценочных шкал целесообразно выражать отклонение не в абсолютных величинах, а в процентах, где за 100% берется величина фонового состояния. При оценке нельзя упускать из виду, в какую сторону идет отклонение от фона в экологическом и физиологическом смысле. В экстремальных случаях наблюдаемся отклонение и в положительную сторону. В конечном итоге получается набор величин, количество и величина отклонений которых отражает состояние экосистемы. Чем больше отличие от фона и чем выше отклонение, тем «хуже» состояние экосистем. В зависимости от распределения отклонений по уровням индикаторов (популяций и индивидов) можно судить о степени и характере изменеиий состояния экосистем (рисунок).
Рисунок — Модель оценочной шкалы
Резкое отклонение физиологических индикационных признаков и почти фоновое значение экологических свидетельствуют о ранних стадиях нарушения экосистем. На стадии структурного нарушения экосистем экологические и физиологические признаки достигают почти одинаковых отклонений и они пропорциональны. О функциональном нарушении экосистем свидетельствует превышение отклонений экологических признаков над физиологическими. На этом этапе физиологические признаки уже не меняются, более качественная оценка получается за счет экологических признаков.
Система оптимально подобранных биотестов позволяет получить наиболее адекватную информацию о степени опасности различного типа техногенных загрязнителей.
На первом этапе на основании фондового и катрографического материала создается карта в масштабе, соответствующем задачам мониторинга. Проводится ландшафтное, геоботаническое и почвенное картирование района с выделением коренных стабильных биоценозов, деградирующих в результате интенсивной антропогенной деятельности, и биоценозов, находящихся на разных стадиях восстановительной сукцессии.Следующий этап – выделение видов индикаторов, изменение в реакциях которых на поведенческом или аналогичном уровне очевидно и интенсивность которого может быть определена достаточно быстро с помощью экспресс-методов: билатеральная асимметрия, ловчие сети пауков-крестовиков, тератогенез растений, видовое разнообразие птиц, животных и т.д. В зависимости от результатов предварительных исследований планируется необходимое количество биотестов, позволяющих получить систему динамических показателей, характеризующих состояние основных сред биоценозов: воздушную, наземную, почвенную и водную. Независимо от природы антропогенных факторов биотесты качества окружающей среды должны оценивать устойчивость биоценозов, популяций, видовое разнообразие, интенсивность размножения и динамику ростовых процессов.Для оценки степени канцерогенности воздействующих факторов можно использовать частоту встречаемости онкологических заболеваний у различных групп животных и в зависимости от особенностей их обитания провести дополнительные исследования.
Рассмотрим последовательность проведения процесса моделирования по биомониторингу природной среды. В зависимости от цели мониторинга, типов загрязнения природной среды, финансовых затрат и научного потенциала коллектива определяется размер обследуемой территории. На основании ландшафтных карт и геоботанической информации определяются типы биоценозов. Проводится ранжирование биоценозов по степени их антропогенной нарушенности в рамках концепции регрессионной или восстановительной сукцессии. Выявляются доминантные, эдификаторные и исчезающие виды растений и животных. Определяется необходимый и по возможности достаточный комплекс биотестов, позволяющий оценить состояние атмосферы, гидросферы и литосферы исследуемого района в зависимости от типа антропогенных загрязнений и возможности рекультивационных мероприятий.
Определяются виды – указатели и индикаторы степени антропогенных загрязнений, с последующим определением дозы (зависимой кривой) в экспериментальных условиях. Определяется временная последовательность при использовании биотестов (сутки, месяц, год…). Определяется площадь элементарной анализируемой пространственной ячейки.Пространственное размещение биотестов с учетом размерности каждого биотеста и площади элементарной пространственной ячейки, с целью компактного заполнения информацией всей последовательности анализируемого пространства. Производится разделение мониторингового района по выбранной системе элементарных пространственных ячеек и заполнение их информацией биотестов. Для токсикантов, способных накапливаться в трофических цепях биоценозов: радионуклеиды, тяжелые металлы и т.д., определяются коэффициенты накопления в каждом трофическом уровне в зависимости от типа биоценоза.
Оцениваются коэффициенты миграции токсикантов при биологическом переносе веществ основными группами животных. Определяются коэффициенты битрансформации загрязняющих веществ, обусловливающих повышение их токсичности в результате биохимических реакций при миграции в трофических цепях (ртуть — метилртуть).Осуществляется предварительная обработка получаемых результатов с представлением их в виде безразмерных единиц и экспертной оценкой специалистов: количественной в единой системе баллов, качественной: хорошо, плохо, удовлетворительно. Проводится статистическая обработка пространственно временной структуры данных биотестов с целью текущей оценки ситуации. Разрабатываются различного типа прогностические модели на основании полученной информации с целью вероятностного прогноза последующего развития экологической ситуации. Экспериментальные исследования: лабораторные и полевые с целью выяснения зависимости доза-биологическая реакция для отдельных антропогенных факторов при их комплексном воздействии, характерном для данной местности, например: тяжелые металлы, бифенолы, электромагнитные поля, радиация.Экспериментальные исследования проводятся дифференцированно для биосистем различного уровня организации: микроорганизмов, растений, беспозвоночных, рыб, амфибий, пресмыкающихся, птиц, млекопитающих и человека. В качестве объектов исследования используются, как правило, потенциальные виды-индикаторы.Первый уровень чувствительности биообъектов к влиянию техногенных факторов проводится на рефлекторном или поведенческом уровне.
Среди растений хорошим объектом исследования является, например, мимоза, которая сокращает поверхность листовой пластинки при действии неблагопритных факторов. Среди беспозвоночных – пауки-кругопряды, дождевые черви, которых можно вместе с почвой помещать между двумя стеклами, где они живут и перемещаются по своим ходам. При добавлении расчетной концентрации загрязняющих почву веществ происходит изменение их двигательной реакции, которую можно наблюдать визуально. Отличным объектом также являются аквариумные и речные рыбы и моллюски. Для млекопитающих разработано достаточно большое количество поведенческих лабораторных методик, позволяющих наблюдать за изменениями в их поведении при воздействии раздражающих факторов.
Вторая, наиболее чувствительная группа тестов – эмбриотропные тесты, с помощью которых возможно оценить степень влияния токсикантов на эмбриональное развитие – самый чувствительный период индивидуального развития организма. Для растений – всхожесть семян, для беспозвоночных – личиночные стадии онтогенеза, для рыб и амфибий – развитие икры, рептилии и птицы – период до выклевывания из яйца, млекопитающие – период беременности. При этом учитывается количество появившихся потомков по отношению к числу зародышей, типы аномалий развития и число особей, доживших до половозрелого возраста, пол, вес и размеры.
В связи с тем, что современные техногенные факторы в последние 40 лет приобрели хронический характер, необходимо проводить специальные исследования по хроническому влиянию загрязнителей в течении 70-80% периода онтогенеза живых организмов. Начиная с рождения, подобные биообъекты должны в экспериментальных условиях и при наличии параллельного контроля подвергаться постоянному хроническому влиянию исследуемого фактора, или совокупности доминирующих загрязнителей до старения, либо до гибели 50% подопытных объектов.
Только при хроническом влиянии факторов возможно получение достаточно полной информации с учетом критических периодов постнатального онтогенеза, в которых устойчивость организма может снижаться в несколько раз.В качестве тест-реакций необходимо использовать основные биохимические и физиологические показатели, видоспецифичные для каждой группы биосистем.Последующий этап включает в себя изучение изучение отрицательного влияния антропогенных факторов в ряду поколений животных. При хроническом действии факторов скрещиваются потомства в течении 10-20 поколений. Отмечают отход животных в разных возрастных группах, уродства, типы патологий и проводят генетический контроль в соматических и половых клетках животных.Как показали исследования по влиянию слабых доз радиации, наиболее выраженные отрицательные последствия в популяции лабораторных мышей наблюдались в первом, третьем, седьмом и двенадцатом поколении животных.Следует также отметить, что, наряду с лабораторными экспериментальными исследованиями,достаточно часто используются эксперименты в естественных условиях.Индикаторные биообъекты помещают в загрязненные природные условия с известными количественными характеристиками воздействующих факторов и с определенной периодичностью фиксируют необходимый комплекс реакций, который сравнивается с показателями контрольной группы.После анализа всей совокупности экспериментальных данных и наблюдений в природе выбирается группа биоиндикаторов, которая рекомендуется для использования в системе того или иного типа мониторинга.Таким образом, научно обоснованный экологический мониторинг позволяет в динамике прослеживать степень антропогенного влияния на природную среду. Динамические ряды наблюдения по опробированной системе биотестов позволяют прогнозировать состояние исследуемых экосистем, разрабатывать оптимальные природоохранные мероприятия и обосновывать необходимые экологические санкции.При отборе видов следует учитывать их пространственное распределение, т.к. те виды, которые имеют небольшую плотность, менее удобны для мониторинга. Идеальными для моделирования на популяционном уровне являются чувствительные, ключевые виды. Подвижность ключевого вида должны быть невысокой, чтобы иммиграция и эммиграция не влияли на конечный результат. Предпочтительны виды, ведущие оседлый образ жизни.
У высших животных, включая человека, при всем разнообразии индивидуальных адаптаций развитие её характеризуется общими закономерностями. Выделяются 2 этапа:
1. Первоначальная – несовершенная адаптация.
2. Долговременная совершенная адаптация.
Для обеспечения дополнительных энергетических затрат организма в процессе эволюции сформировался специальный физиологический механизм, названный общим адаптационным синдромом (ОАС).Независимо от типа влияния естественного или антропогенного фактора при условии превышения его физиологического уровня в организме, развивается ОАС.
Например: появление и развитие злокачественных заболеваний используется моделирование в качестве биотеста, указывающего на полное несоответствие окружающей среды адаптационным возможностям организма. В качестве примера рассмотрим ситуацию с судаком, запущенным в 1963 – 1964 гг в качестве промысловой рыбы в озеро Балхаш, на берегу которого находился один из крупнейших в бывшем СССР горно–металлургических комбинатов, сливавший все свои неочищенные отходы в озеро. Выпущенный судак интенсивно размножился. Являясь хищником, активно стал накапливать канцерогенные вещества. В 1969 году наблюдалось массовое поражение рыбы злокачественными образованиями. В дальнейшем наблюдалась массовая гибель рыбы.Различные органы по-разному накапливают радионуклиды, тяжелые металлы и токсические соединения: стронций и цезий активно накапливаются в костях животных, в печени и в почках содержание радионуклидов превышает их содержание в мышцах в 1000 раз. Метил -ртутные соединения вызывают нейротоксические эффекты.
Комбинацией разных фито- и эооиндикаторов на двух системных уровнях можно оценить постепенные и очень мелкие изменения в состоянии экосистем. И при четком наборе характерных для изучаемой территории ключевых участков можно оценить и более крупные географические единицы и составить карту состояния окружающей среды. На основе карты и оценочных шкал можно в будущем очень быстро оценить состояние любой экосистемы соответствующего типа данного региона или при наличии фоновых экосистем и других регионов и дать рекомендации обоптимальной хозяйственной деятельности на данной территории.
Контрольные вопросы
1. Отбор показательых видов на популяционном уровне биоиндикации
2. Показатели популяционного уровня биоиндикации.
3. Воздействие антропогенных стрессоров на динамику растительных популяций
4. Воздействие антропогенных стрессоров на характер распространения растений
5. Показательные признаки экосистемного уровня
6. Метод комплексной биоиндикации, его этапы и преимущества
7. Экологические индексы, используемые в методе комплексной индикации (индекс Шеннона, идекс домиантности, индекс сходства)
Достарыңызбен бөлісу: